1 I primati, Distribuzione e origini
Per approfondire: Primati
Slide : Seconda lezione, link
I primati non umani attualmente presenti sono distribuiti entro i due tropici in tutti i continenti eccetto l'Oceania. Unica eccezione sono delle specie presenti in Marocco (macaco) e nell'Arcipelago giapponese. Approfondisci qui
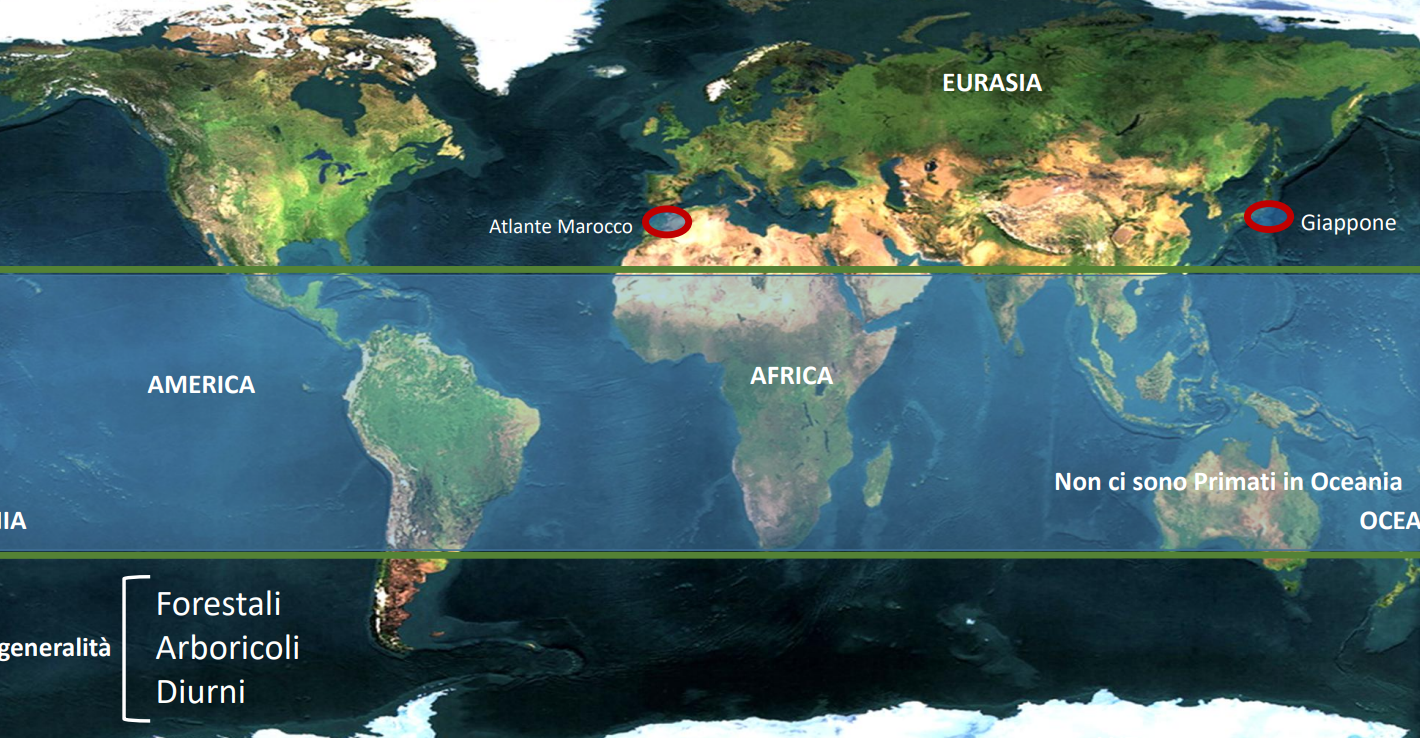
Generalmente sono animali tipici di ambienti forestali, con specializzazioni arboricole e diurne (con delle eccezioni).
L'adattamento ha uno stile di vita arboricolo ha portato a diverse specializzazioni pre-adattative ad alcune innovazioni umane. Approfondisci qui
Sistematica: Approfondisci qui
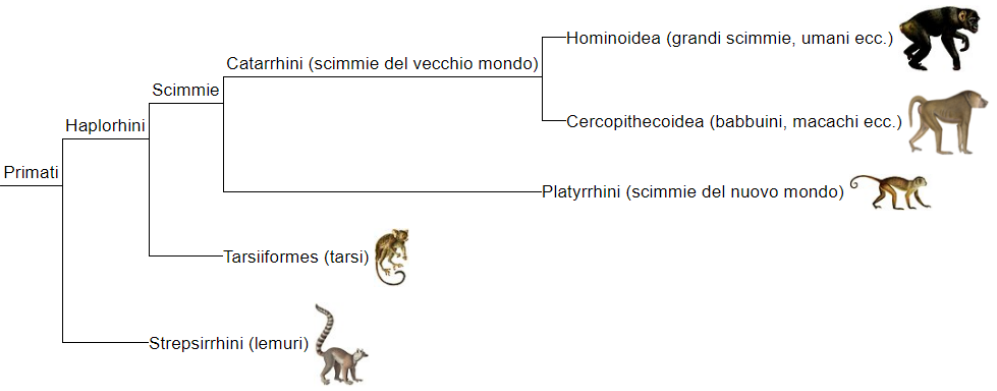
Discriminante nella distinzione tra Haplorrhini e Strepsirrhini è la presenza o assenza del rinario. I Tarsiformi sono un gruppo particolare con molti caratteri unici, non facilmente collocabili. (spesso sono inseriti entro gli Haplorrhini, il libro mi sembra li separi)
All'interno dei Simiiformes si possono distinguere i due gruppi su base biogeografica. I Platyrrhini sono le scimmie del nuovo mondo, mentre i Catarrhini sono scimmie del vecchio mondo. Ci sono anche altre caratteristiche che le distinguono, come la direzione del rinario (verso il basso per le Catarrine, laterale per le Platyrrhine), la formula dentale. Ci sono anche tendenze locomotorie e dietetiche diverse.
Le Catarrine sono distinte in Cercopithecoidea che contiene specie come babbuini e macachi, diffusi in Africa, Giappone, Gibilterra Asia SudOrientale; e i Hominoidea, in un'areale più ristretto (Africa Centrale e SudEst Asiatico).
Gli Hominoidea sono ulteriormente divisi in sottogruppi:
Origini! (Non nelle slide ma nel libro?)
2 Chi sono gli ominoidi attuali
Slide: Seconda lezione, link
Hominoidea è una superfamiglia interna ai Primati.
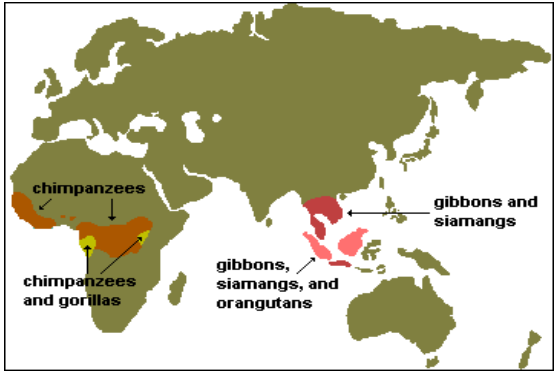
Sono divisi nei seguenti sottogruppi, e distribuiti in Africa Centrale e nel SudEst Asiatico.
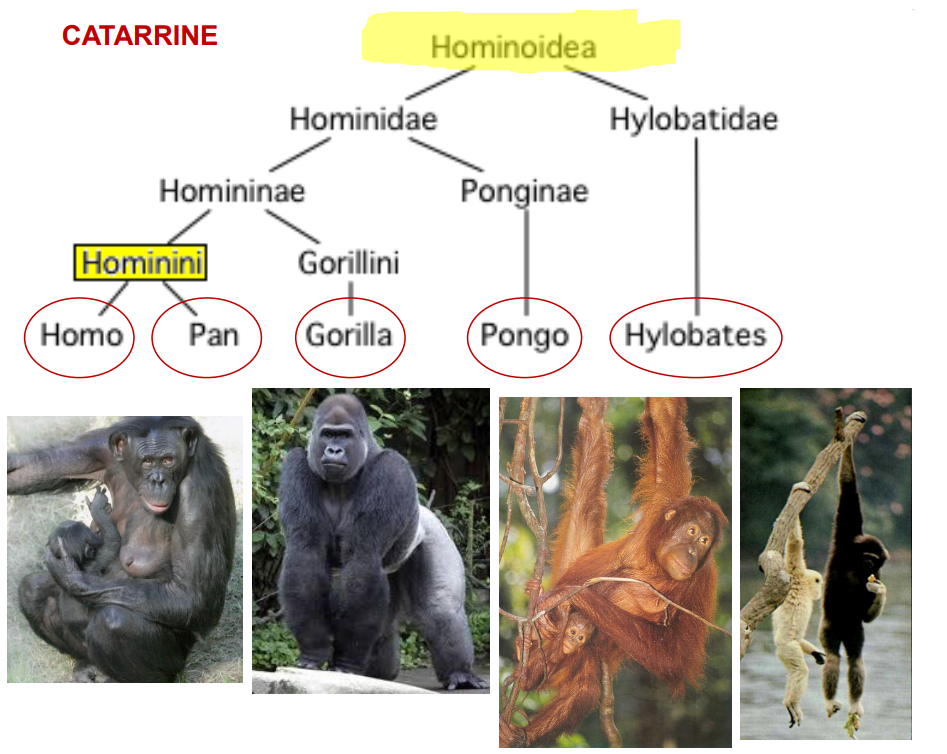
Hylobatidae
Comprendono i gibboni e siamango. Abitano foreste tropicali e sudtropicali dell'Asia sudorientale, in particolare Giava, Borneo e Sumatra.
Sono brachiatori, divisi in coppie monogame e scarso dimorfismo sessuale.

Ponginae
Parte della famiglia Hominidae.

Orangutan (Pongo pygmaeus) è l’unica specie dei Pongidi, vive nelle foreste del Borneo (P. p. abelii) e di Sumatra (P.p. pygmaeus) in Indonesia.
Gorilla - Gorilla gorilla
Parte di Homininae.

La più grande delle scimmie antropomorfe, con un forte dimorfismo sessuale. La locomozione è quadrupede. Distribuito in Africa Centrale.
Pan spp.
Divisi in Pan paniscus (bonobo) nella RDC e Pan troglodytes (scimpanzé), Sierra Leone e Guinea fino ai laghi Tanganica e Vittoria.
Con un vasto repertorio motorio, scarso dimorfismo sessuale. Sia arboricoli che terrestri.

Hanno comunità complesse, caratterizzati da poliginia. C'è una mobilità delle femmine e una scarsa mobilità dei maschi. Vegetariani al 90%, i maschi sono più aggressivi e organizzano battute di caccia.
Uso di utensili la cui fabbricazione e uso sono appresi.
3 Cambiamenti climatici nel terziario ed evoluzione degli ominini
Slide: terza lezione, link
Approfondisci qui
Nell’Oligocene e inizio Miocene le comunicazioni tra Europa e Africa sono interrotte.
Tra circa 30 e 22 Ma fa c'è una lacuna di testimonianze di fossili africani (African Ape Gape – AAG).
Nell'Oligocene cominciano a diversificarsi i primi animali ora attribuibili al clado dei Siimiformes. Approfondisci
A partire dal Miocene inferiore (circa 20- 16 Ma) in Kenya e in Uganda si ritrovano scimmie con caratteristiche ominoidi.
Proconsul è un importante ominoide miocenico evolutosi in Africa. Approfondisci
Nel Miocene c'è un importante scambio faunistico tra Africa e Eurasia, grazia a un contatto diretto tra i due continenti. (Miocene medio, 17Ma). Approfondisci
Discendenti di Proconsul si diffondono in Eurasia e diversificano. Si rinviene Oreopithecus in Sardegna, con forme capaci di brachiazione. In Asia Sudorientale si rinviene un cluster di scimmie antropomorfe che daranno probabilmente origine ai Orangotanghi attuali.
nel Miocene superiore avviene la crisi di salinità del Messiniano (6Ma).
In Africa si incontrano nuove forme di Ominini, come Orririn sp. Ardipithecus sp. che presentano un mosaico di strutture simili a quelli delle scimmie del genere Pan attuali, con postura eretta e bipedismo facoltativo (non specializzato tanto quanto gli Australopitechi). Approfondisci in Hominini
In generale il bipedismo viene sperimentato a partire da 6 milioni di anni fa da specie che vivono in ambienti misti. Dai 3 milioni di anni fa diventa molto importante per specie costrette a vivere nei nuovi ambienti di savana.
A partire da 4 milioni di anni fa, i reperti cominciano ad essere più numerosi.
Nell'epoca Pliocenica e parte del Pleistocene inferiore sono state descritte 7 specie di Ominini, per la maggior parte attribuite ad Australopithecus.
Sono un gruppo di specie i cui fossili sono stati ritrovati in varie zone dell'Africa orientale, meridionale e centrale.
Aumentano le dimensioni dentarie e hanno una palese andatura bipede detta facoltativa. Mantenevano la capacità di arrampicarsi per avere un accesso maggiore a risorse diversificate. Questa diversificazione del repertorio locomotorio era probabilmente una risposta all'inaridimento del clima africano.
Con la comparsa di Australopithecus e di Paranthropus si specializza ulteriormente il bipedismo, data la comparsa degli ambienti di Savana. Le prime specie di Australopithecus compaiono dai 4,2 Ma nel Pliocene, mentre le prime specie di Paranthropus compaiono dai 2,7Ma a partire da Pleistocene. Approfondisci
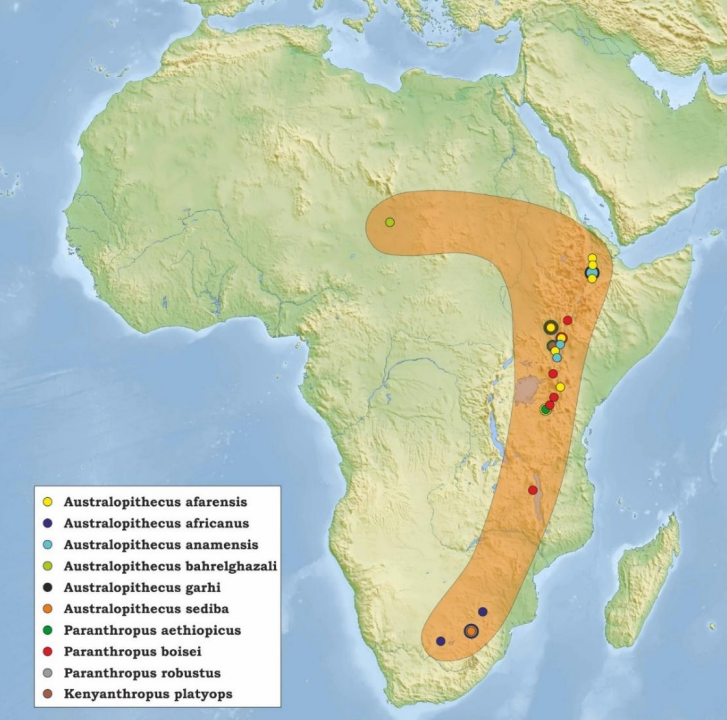
I primi Australopitechi (A. afarensis) non erano particolarmente specializzati (sono comunque Ominini com locomozione più derivata di Ardipithecus). La bipedia restava facoltativa e mantennero una buona capacità di arrampicamento. Due reperti importanti sono Lucy e le impronte di Laetoli.
Potrebbe aver utilizzato Approfondisci.
C'era un forte dimorfismo sessuale, si deduce che la organizzazione sociale potesse essere simile agli attuali gorilla. Approfondisci
Con A. africanus, di taglia maggiore rispetto ad A. afarensis. il bipedismo diventa obbligato. Approfondisci
Il Terziario
Chiamato anche Cenozoico (Nuova vita), ha avuto inizio 65,5 Ma ed è tutt'ora in corso. Il Quaternario è considerato al suo interno.
A livello climatico, soprattutto a partire dal Miocene (23-5Ma) si assiste a ampie oscillazioni di temperatura e umidità, con l'alternarsi di periodi aridi a periodi umidi.
| xx | |
|---|---|
| Oligocene | 34 - 23 Ma |
| Miocene | 23 - 5,3 Ma |
| Pliocene | 5,3 - 2,6 Ma |
| Pleistocene | 2,6 - 0,0 12Ma |
| Wikipedia: |
4 Cambiamenti climatici nel quaternario ed evoluzione degli ominini
Slide: terza lezione, link
Per approfondire questi argomenti Qui e Anche qui
Nel Quaternario ancora assistiamo all'evoluzione di alcune specie di Australopitechi. Tra queste vediamo A. sediba, rinvenuto in SudAfrica Approfondisci
Si può notare che in questi ominini c'è una riorganizzazione della regione orbitofrontale, che indica uno sviluppo simile a quello del genere Homo. Questa riorganizzazione deve precedere lo sviluppo della capacità cerebrale.
Inoltre, la conformazione del bacino è più simile a quella dell'uomo moderno, ma più per esigenze locomotorie (bipedismo) che per esigenze ostetriche, siccome la capacità cranica non era ancora troppo grande.
Nel passaggio tra Pliocene e Pleistocene si aridificano le regioni africane prima caratterizzate da foreste dense, in cui si sono evolute diverse forme di scimmie. Questo processo di aridificazione ha portato a una progressiva comparsa delle savane. Questo nuovo ambiente ha richiesto nuovi adattamenti e ha consentito la comparsa di Ominini come Paranthropus (con diete specializzate su radici e piante erbacee C4, cioè "cibi di ripiego"). Anche il bipedismo è associato a questo nuovo ambiente, già apparso precedentemente, seppur non molto specializzato, è stato cooptato da diverse specie in forme più specializzate, che hanno perso progressivamente le capacità arboricole. (A. africanus e sediba, Homo)
Concomitanti ai Australopitechi e alle prime specie attribuibili al genere Homo, nel Pleistocene vediamo l'apparizione di scimmie del genere Paranthropus, più robusta, con un apparato masticatorio molto accentuato e specializzato, ma che scompariranno 1Ma.
Caratterizzante del genere è la presenza di una cresta saggittale e un apparato masticatorio molto accentuato, indice di una dieta specializzata (più specializzata in P. boisei che P. robustus). Approfondisci
In questo periodo inoltre compaiono le prime tecnologie litiche, inizia quindi il Paleolitico o ESA (Early Stone Age). Dai 3,3 Ma in Kenya a possibili tracce di utilizzo in Etiopia, attribuite a A. afarensis. Altre tecnologie sono state retrodatate ai 3-2,5Ma e attribuite a H. habilis e Paranthropus.
Approfondisci
In questo periodo questi ominini cominciano a spostarsi dall'Africa orientale anche all'Africa settentrionale (reperti rinvenuti in Algeria).
Il Quaternario
Chiamato anche Neozoico, ha avuto inizio 2,5Ma ed è tutt'ora in corso. E' l'ultimo periodo del Terziario (o Cenozoico).
E' stato un periodo caratterizzato da forti oscillazioni di temperatura e umidità, è un periodo che è stato attraversato da sette ere glaciali.
Inoltre nel passaggio da Pliocene a Pleistocene (quindi da terziario a quaternario) avviene la comparsa delle Savane, congruente con l'evoluzione dei generi di Paranthropus e Homo.
Nel Pleistocene inferiore si assiste a una graduale scomparsa di bipedi del genere Paranthropus e Australopithecus e la nascita e affermazione del genere Homo
Nel Pleistocene medio c'è un'elevata variabilità morfologica del genere Homo in aree geografiche distinte (Africa, Asia, Europa). Evolvono e appaiono altre specie, come H. neanderthalensis e H. sapiens
Nel Pleistocene superiore scompaiono molte specie del genere Homo e si afferma H. sapiens, che arriva a colonizzare il continente americano.
| Pleistocene inferiore | 2,6 Ma - 770 Ka | Raffreddamento delle temperature |
|---|---|---|
| Pleistocene medio | 770 - 126 Ka | Inversione campo magnetico |
| Pleistocene superiore | 126 - 12 Ka | |
| Olocene | 12 Ka - attuale |
5 Il Bipedismo, quando compare, in che specie e con quali caratteristiche
Slide: terza lezione, link
Homo sapiens ha diverse caratteristiche peculiari, che si ritrovano nell'evoluzione dei primati.
- Postura eretta obbligata (bipedismo inizia col raddrizzamento del tronco).
- Capacità cranica elevata (dai 3 Ma)
- Faccia molto piccola rispetto al cranio (dai 300'000 anni fa, e globularità del cranio)
- Mano libera
Il bipedismo appare dai 7 Ma con gli Homini appartenenti ai generi Sahelanthropus, del quale è stato rinvenuto un cranio con un foro avanzato, indicativo di una postura eretta, ma non abituale, e del genere Orririn, di cui è stato rinvenuto un femore che potrebbe indicare una andatura bipede di tipo facoltativo. Approfondisci
poi in Ardipithecus (4,4 Ma) vediamo sempre caratteri indicativi di un bipedismo primitivo, tra cui un foro occipitale in posizione avanzata che indica una posizione eretta; (comunque meno specializzato degli Australopitechi) seppur permane una capacità di arrampicamento, data dai alluci opponibili (ma non di brachiazione e sospensione). Approfondisci
La postura eretta e l'andatura bipede potrebbero essere evolute più volte indipendentemente. Inoltre, dato che l'ambiente in cui queste forme antiche era probabilmente forestale, è probabile che la postura eretta e il bipedismo fossero un preadattamento al tipo di locomozione più attestato nei nostri antenati più recenti. Non necessariamente è nato come una risposta all'adattamento a un ambiente di savana aperto, ma è stato cooptato quando quest'ambiente è apparso.
Nei Australopitechi la andatura bipede diventa più specializzata (pollice parallelo al piano del piede), seppur permane una certa capacità di arrampicamento. Le forme più antiche (come A. afarensis) erano meno specializzate, mentre le forme più recenti (A. africanus, A. sediba) mostrano un bipedismo più specializzato e minori capacità di arrampicamento, siccome passano da ambienti forestali marginali alla savana ad ambienti di savana e boscaglia.
Approfondisci
Un esempio è lo scheletro postcraniale attribuito a A. afarensis e noto come "Lucy", mostra diversi adattamenti utili a una postura bipede facoltativa:
Un esempio è l'alluce addotto (parallelo alle altre dita) e non più divergente o il femore inclinato rispetto alla tibia. Tuttavia permangono molti caratteri che denotano una capacità di arrampicamento, come le braccia allungate.
In A. africanus pare che il bipedismo fosse già obbligato. (lo dice nelle slide ma non lo supporta).
In A. sediba si vede una evoluzione della morfologia del bacino, per accogliere una locomozione più specializzata.
7 Gli Ominini pliocenici (Sahelanthropus, Orrorin, Ardipithecus, prime forme di Australopithecus)
Vabbe ne ho già parlato un sacco boiaeva
Approfondisci tutto qua
Slide: terza lezione, link
Sahelanthropus tchadensis
E' stato rinvenuto nel deserto del Chad, in Africa centrale. E' datato a circa 7 Ma. (Miocene superiore)
Alcune caratteristiche peculiari sono una posizione avanzata del foro occipatale, che indica una possibile postura eretta. Restano però caratteristiche "arcaiche", come una taglia corporea e cerebrale simile a quella dei Scimpanzè attuali.
L'andamento bipede però non era abituale.
Orrorin tugenensis
Un insieme di fossili è stato rinvenuto in Kenya e datati intorno al 6 Ma (verso la fine del Miocene) e attribuiti a Orrorin tugenensis. Di questi dei frammenti di femori sembrano indivare una capacità di postura eretta e andatura bipede, però diversa da quella documentata in specie più recenti.
In entrambe le specie c'era probabilmente uno scarso dimorfismo sessuale.
Approfondimento Sahelanthropus e Orrorin
Ardipithecus ramidus
Sono stati rinvenuti reperti appartenenti a circa 36 individui, in un ambiente ancora forestale, datati a 4,4 Ma (tra Miocene e Pliocene)
Il dimorfismo sessuale era scarso (minore che Australopithecus). L'andatura bipede era ancora facoltativa, seppure erano assenti indicatori di abilità brachiatoria e sospensione.
Approfondisci
Australopithecus afarensis, anamensis
Accorpati perchè mostrano morfologie sostanzialmente simili. A. anamensis è più antico
- Taglia corporea e cerebrale tipo scimpanzé
- Denti a morfologia variabile (per morfologia e spessore dello smalto) – diastema, I molari sono più grandi e lo smalto è più spesso di Ardipithecus
- Prognatismo
- Capacità cranica 300-400 cc
- Elevato dimorfismo sessuale (M: 130 cm, 45 kg; F: 100 cm, 30 kg)
- Locomozione mista: capacità arrampicamento e bipedia (ma non obbligata)
Approfondisci
Reperti importanti sono quello rinominato come "Lucy", datato 3,2 Ma e rinvenuto ad Hadar, in Etiopia, attribuito a A. afarensis, che mostra le caratteristiche sopraelencate.
Un altro reperto importante attribuito a A. afarensis sono le impronte di Laetoli trovate in Tanzania e datate a 3.6 Ma, che mostrano l'alluce addotto.
Erano specie ad elevato dimorfismo sessuale, con una struttura sociale ipoteticamente simila a quella dei Gorilla attuali. Approfondisci
8 Gli Ominini del Pleistocene (Australopithecus, Paranthropus, Homo e le diverse specie)
Slide: terza lezione, link e quarta lezione (Homo), link
Nel Pleistocene si assiste a una aridicazione dell'Africa, con la comparsa della Savana. Diminuisce quindi l'habitat forestale e molte forme che già cooptavano il bipedismo sono costrette a specializzarlo.
Australopithecus africanus
Tra queste ci sono nuove forme del genere Australopithecus, come A. africanus.
Un reperto importante attribuito a questa specie è il bambino di Tuang, rinvenuto in SudAfrica, datato a 2,3 Ma, che mostra una velocità di crescita maggiore rispetto all'uomo moderno. Approfondisci
Australopithecus sediba
Rinvenuti in SudAfrica, due scheletri parziali datati tra gli 1.9 e 1.7 Ma.
Mostrano apomorfie con il genere Homo, come il pollice lungo e l'accorciamento delle altre dita (per consentire una presa di precisione). Approfondisci
La completezza dei reperti ci fornisce molte informazioni sulle innovazioni evolutive di questa linea di ominini, coerenti con il successivo sviluppo umano:
- Riorganizzazione e aumento del cervello
- Riduzione dei denti
- Sporgenza delle ossa nasali (termoregolazione)
- Aumento della taglia corporea
- Riorganizzazione biomeccanica della pelvi in relazione alla locomozione
- Relativo allungamento degli arti inferiori
- Aumento dell’arco longitudinale del piede
- Potenziale uso e fabbricazione di strumenti
In questo periodo appare anche un nuovo genere di ominini, chiamato Paranthropus
Caratteristiche generali del genere sono una particolare morfologia dell'apparato masticatore, con denti laterali più grandi a scapito di quelli anteriori, e uno smalto dentario molto spesso. Ciò consentiva in alcune specie del genere una dieta specializzata.
Molti crani attribuiti al genere inoltre presentano una cresta saggittale. Approfondisci
Il genere si estinguirà attorno a 1 Ma, probabilmente perchè la dieta specializzata non riesce a far fronte ai cambiamenti climatici del tempo.
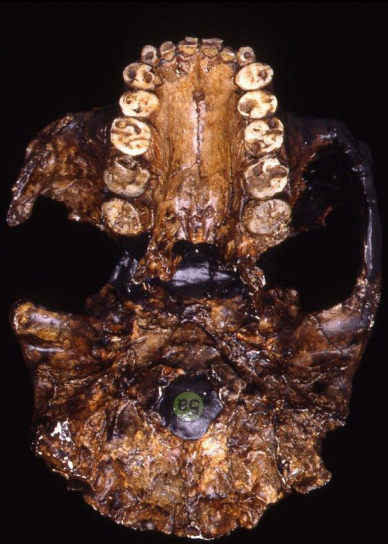
Paranthropus aethiopicus
Un cranio datato a 2,5 milioni di anni fa, rinvenuto in Kenya. Mostra caratteristiche generali del genere (apparato dentale grande e specializzato, cresta saggitale) ed una capacità cranica di circa 410cc.
Paranthropus boisei
Qui la specializzazione dell'apparato dentario è indice della possibile dieta della specie. Dato un ambiente con una disponibilità limitata di varietà di risorse, la specie ha denti laterali molto specializzati per il consumo di piante erbacee di tipo C4 e radici. Approfondisci
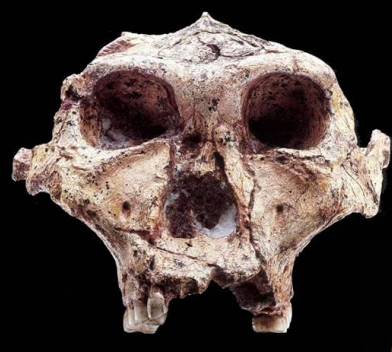
Paranthropus robustus
Probabilmente la dieta era meno specializzata di P. boisei, e la specie faceva ricorso a un ampio spettro di risorse alimentari, con una dieta più simile a quella dei Australopitechi. Approfondisci
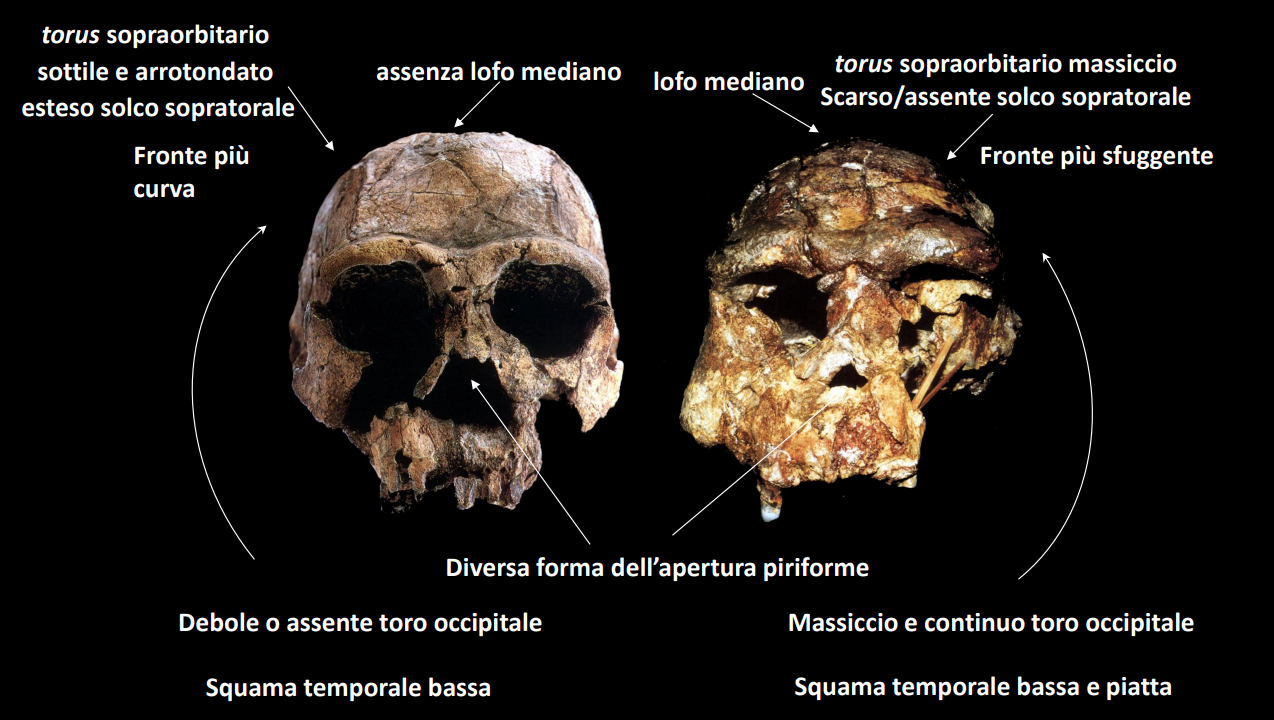
Appare inoltre il genere Homo nelle sue prime forme più primitive. Delle caratteristiche da subito innovative del genere sono un aumento della taglia corporea e delle dimensioni cerebrali. Diminuiscono inoltre i denti posteriori. Approfondisci
All'apparizione del genere sono anche attribuite le prime industrie litiche Leggi qui dhai
Si può associare l'apparizione delle prime tecnologie alle prime industrie e innovazioni portate dal genere, assieme all'introduzione dei nuovi alimenti. Approfondisci
Homo habilis
Il range dei reperti attribuiti alla specie è collocato nell'intervallo tra i 2,8 e 1,4 Ma. Da Koobi Fora. Approfondisci
Homo rudolfensis
Rinvenuto anche questo a Koobi Fora e datato tra i 2-1,8 Ma.
E' interessante che la capacità cranica fosse maggiore rispetto a H. habilis, il prognatismo inoltre era ridotto e i denti laterali avevano dimensioni maggiori, indicando possibilmente una dieta distinta. Approfondisci
Homo ergaster (H. erectus s.l)
Alcuni resti, ritrovati in SudAfrica alla cava di Drimolen dimostrano una coesistenza con altri ominini come Australopitechi e Parantropi.
2,58 Ma - 700 Ka.
E' originata da H. habilis in seguito a eventi di speciazione.
Un reperto importante di H. ergaster è il ragazzo del Turkwana, che mostra un chiaro stacco con le morfologie degli antenati, assieme a molte caratteristiche derivate che poi incontreremo anche in H. sapiens.
Un altro reperto rilevante sono le impronte di Ileret, in Kenya, datate a 1,5 Ma e che mostrano già la presenza di un piede anatomicamente moderno, che implica una locomozione bipede molto simile alla nostra.
Introduce anche una tecnologia litica Modo 2
Approfondimento generale su H. erectus s.l.
Con H. erectus s.l. (sensu lato) si intendono sia le sue forme africane, come H. ergaster che le forme eurasiatiche, indicate come H. erectus s.s. (sensu stricto)
Siccome la specie s.l. occupa uno intervallo cronologico e spaziale molto ampio (dai 2 Ma ai 100 Ka, per tutto il pleistocene), si incontrano morfologie molto distinte nelle forme asiatiche (H. erectus s.s) e africane (H. ergaster)
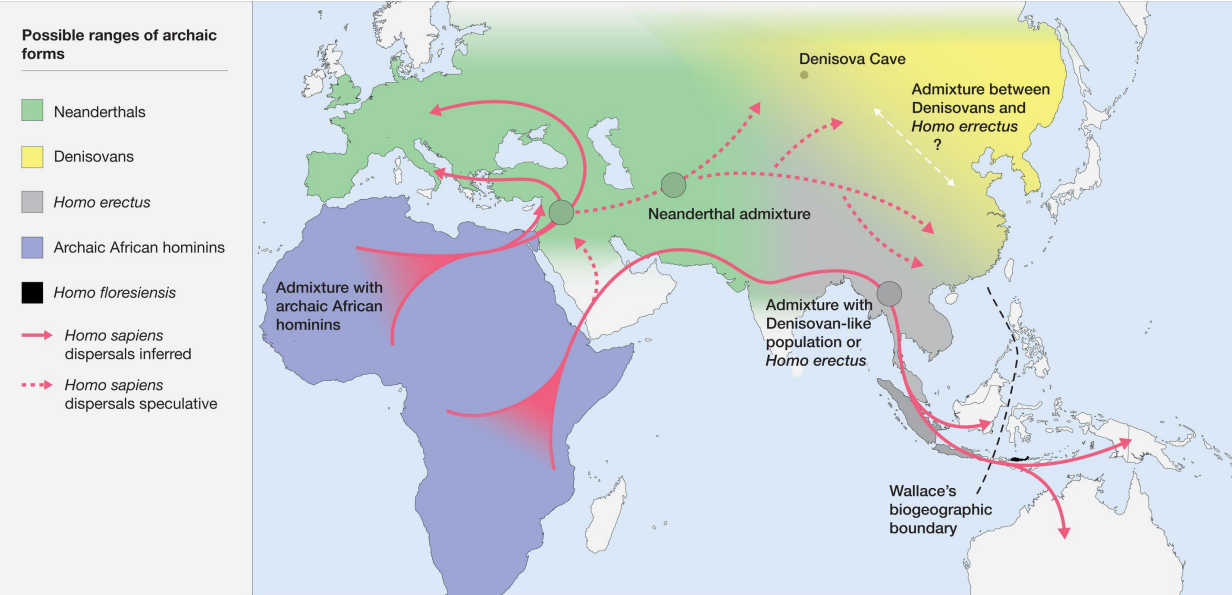
6 I modelli OOA, Quanti ne conosci e a chi vengono associati
Slide: Quarta lezione OOA1, link
Sesta lezione OOA2 in Europa, link
Settima lezione OOA 2 in Asia, link
Undicesima lezione OOA 3 nel Pleistocene medio, link
Dodicesima lezione OOA 3 nel Pleistocene superiore, link
Out of Africa per un approfondimento generale
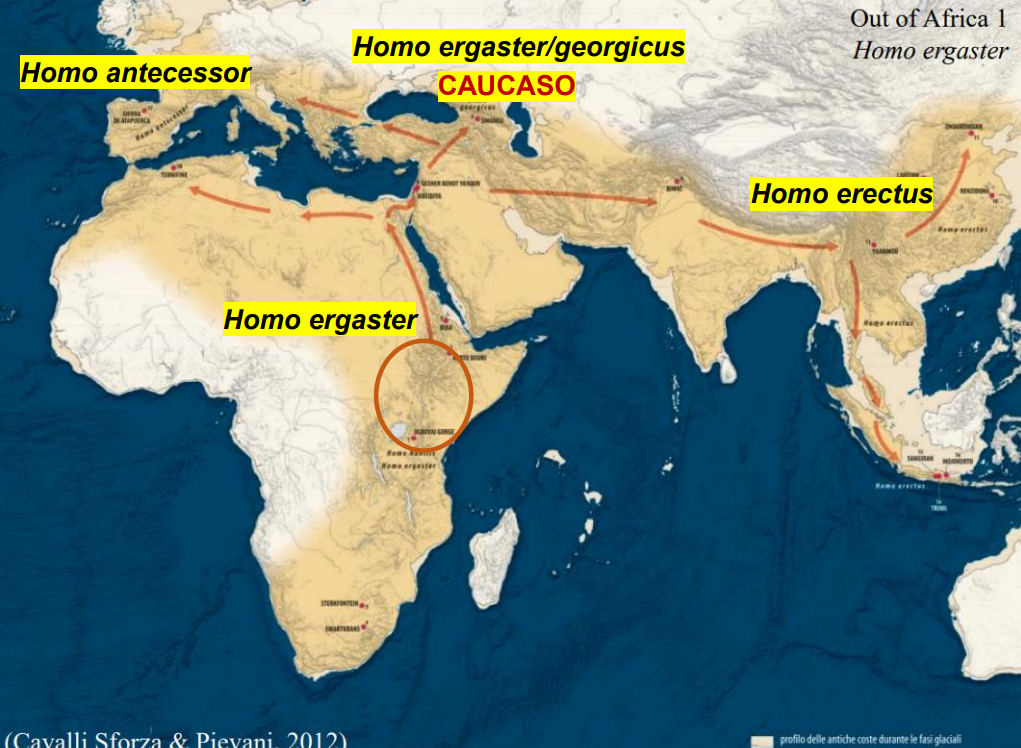
OOA 1
Il primo modello OOA è attribuito a H. ergaster (o H. erectus s.l.).
I primi reperti al di fuori dell'Africa attribuiti a questa specie sono stati rinvenuti in Caucaso, e datati a 1,77 Ma (Dmanisi). Assieme ai reperti sono state trovate industrie litiche Modo 1, ciò suggerisce che si trattasse di popolazioni più arcaiche. Appr.
Successivamente la specie si è spostata in Asia, dove si parla di Homo erectus s.s., ed è soppravvisuta in Indonesia fino ai 118-108 Ka, i reperti asiatici mostrano una grande variabilità morfologica. I primi reperti in Asia sono stati trovati in Cina e datati attorno ai 2 - 1,6 Ma. La retrodatazione di alcuni reperti suggerirebbe una fuoriuscita più antica di quanto suggerito dai reperti di Dmanisi.
In Europa si trovano le prime tracce, seppur non reperti diretti, a Pirro Nord, in Puglia, con dei manufatti litici olduvaiani attribuiti al genere Homo, probabilmente H. ergaster. 1,6-1,3 Ma Appr.
Sempre a Sierra de Atapuerca nel sito di Sima del Elefante è stata rinvenuta una mandibola datata 1,2 Ma, ma non è chiara l'attribuzione a una specie del genere Homo.
In Europa la specie si diversifica e vengono trovati reperti attribuiti a una nuova specie, denominata H. antecessor, rinvenuta a Gran Dolina in Spagna e datati tra i 800-850 Ka.
Appr.
OOA 2
Il secondo modello OOA è attribuito a Homo heidelbergensis. Appr.
L'espansione verso l'Eurasia coincide con momenti di clima temperato nel continente: il pleistocene medio è caratterizzato da forti oscillazioni di temperatura e umidità, date da un'espansione dei cicli di Milankovich (che da 41'000 passano ai 100'000). Questi cicli causano una contrazione o un'espansione delle masse glaciali, che arrivano a coprire il continente Eurasiatico.
L'arrivo di H. heidelbergensis nel continente coincide con un primo periodo caldo, così come la successiva espansione e evoluzione delle nuove specie a partire da H. heidelbergensis.
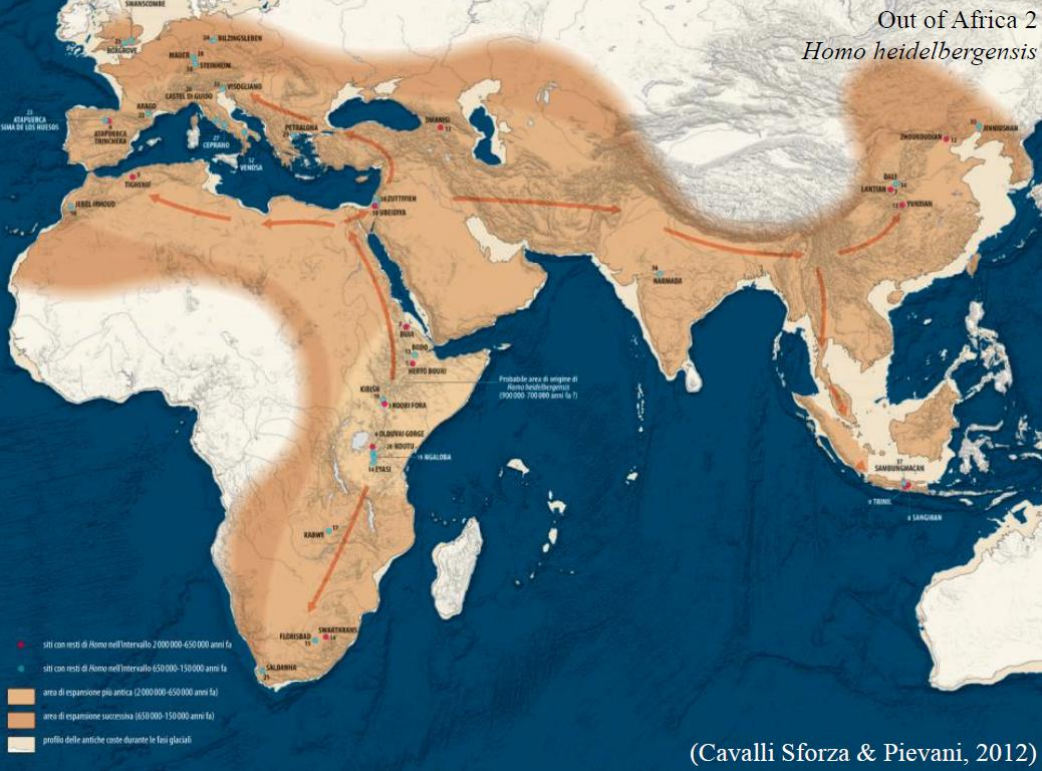
E' importante sottolineare che dall'espansione in Europa di H. heidelbergensis avverrà la graduale comparsa di H. neanderthalensis, in un lasso di tempo che comincia attorno ai 500 Ka. Così come evolvono i Denisoviani, che si differenziano dal resto della linea circa 600 Ka. In Africa emergerà Homo sapiens.
Le prime tracce del popolamento OOA sono resti di bifacciali Modo 2, a Notarchico (Potenza), datate intorno ai 700 Ka. (da checkare, ma mi pare che in eurasia fosse arrivato solo il modo 1). Appr. espansione in Europa
A Ceprano, in Lazio invece troviamo il primo resto attribuito a H. heidelebergensis, si tratta di un cranio parziale, che mostra avere una capacità cranica di 1180-1200 cm^3. E' datato tra i 480-350 Ka.

Si rinvengono altri crani parziali, ad Arago, datato a 450 Ka e Steinheim datato a 250 Ka.
Inoltre è stato rinvenuto in Sud Italia un incisivo mascellare datato tra i 583-561 Ka, appartenente ad un bambino, che potrebbe appartenere ad H. heidelbergensis.
Un sito estremamente importante è Sima de los Huesos, dove i numerosi resti mostrano già un processo di neanderthalizzazione. Inoltre assieme a questi resti si rinviene un industria di Modo 2. I reperti sono datati a 430 Ka.
In Asia non ci sono molti reperti direttamente attribuiti a H. heidelbergensis. Appr.
Data la coesistenza di altre specie (H. erectus, poi i Denisoviani) e caratteristiche che non permettono una attribuzione univoca (molti reperti hanno un misto di caratteri derivati ed arcaici). La datazione è di circa 260 Ka.
Reperti importanti sono il cranio di Jinniushan, e il cranio di Harbin. Non direttamente attribuiti a H. heidelbergensis
OOA 3
La dispersione Out of Africa 3 è attribuita a Homo sapiens.
Il record fossile conferma che è avvenuta a più riprese, sia tramite la Northern route, che tramite il passaggio verso l'Arabia (Southern route), quando il livello del mare lo consentiva (testimoniato da frammenti ritrovati ad Al Wusta, Penisola arabica, datati a 85 Ka)
Il passaggio per il Levante è confermato da diverse testimonianze:
- Mislyia cave, Palestina, 194-177 Ka. Assieme a un frammento di mandibola ci sono tecnologie litiche Levallois, a confermare che questa è stata portata dai Sapiens fuori dall'Africa.
- Quafzeh e Skhul in Palestina, tra i 120-90 Ka. La volta cranica è elevata, con grande capacità (intorno a 1500 cc), la faccia e fronte alte, arcate sopracciliari e sopraorbitarie sviluppate (Skhul). La statura tra 165 e 175 cm. Appr. Quafzeh
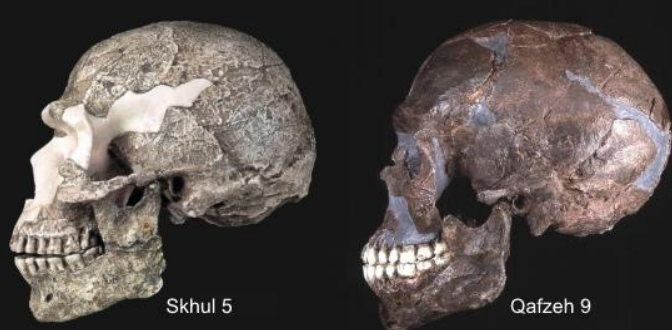
Per dare confronto, la presenza di Neanderthaliani nel Levante non è confermata prima dei 70-60 Ka. Ma il fossile rinvenuto a Manot, Palestina, 55 Ka. pare testimoniare ibridazione con H. n., per via di alcune caratteristiche.
Si sono susseguiti diversi periodi glaciali e interglaciale in questo momento della storia umana. Generalmente, i momenti interglaciali, date le condizioni favorevoli, hanno consentito la migrazione delle popolazioni umane (e quindi l'accesso di Sapiens in Europa), mentre i momenti glaciali hanno frammentato le popolazioni presenti, causando deriva genetica. Appr.
La Grotta Mandrin in Francia testimonia la prima occupazione alternata di Homo sapiens e Neanderthal.
In altri siti, come:
- Bacho Kiro in Bulgaria,
- Zlaty kun in Repubblica Ceca
- Ust'-Ishim in Siberia
si trovano reperti di H. sapiens con chiara introgressione di DNA Neanderthaliano. Tutti questi siti sono datati attorno ai 45 Ka.
Invece, a Grotta del Cavallo (45-43 Ka) in Puglia e Kent (44-41 Ka) in Inghilterra pure si trovano reperti di Sapiens. Queste culture inoltre hanno portato con sé diverse industrie di transizione, innovative per l'Europa.
Tuttavia, nessuno degli individui di H. sapiens arrivati in Europa prima di 40 Ka ha lasciato traccia nelle popolazioni eurasiatiche attuali. Appr.
In Cina vengono trovati dei reperti la cui attribuzione ad Homo sapiens crea dei problemi: essendo datati a 70 Ka Appr., implicherebbe che la specie ha raggiunto prima la Cina che l'Europa, e che ha raggiunto l'Asia ben prima di quanto si pensava Appr.
Forse I neanderthaliani hanno davvero costituito una barriera ecologica all'ingresso di H. sapiens in Europa.
A maggior ragione un serie di reperti rinvenuti a Sumatra e in Australia confermerebbero un arrivo di H. sapiens molto prima che in Europa (70-60 Ka) Appr.
Oltre a tecnologie litiche, a Lake Mungo, Australia (71-51 Ka) si trovano resti parziali di crani dell'essere umano moderno, e possibili segni della prima cremazione confermata. Inoltre, in Borneo (52-40 Ka) si trovano le prime pitture rupestri attestate a H. sapiens
Le popolazioni uscite dall'Africa, non si sono espanse in tutta l'Eurasia fino a circa 45 Ka. La prima tappa è stato un hub, tra Iran, Mesopotamia e Golfo Persico (allora emerso), dove le popolazioni si sono "stanziate" prima di spingersi verso l'Asia e l'Europa. Evidenze genetiche, paleoecologiche e paleoclimatiche supporterebbero quest'ipotesi. Appr.
Il modello demografico è di maggiore successo rispetto a Neanderthal, le popolazioni sono relativamente numerose e interconnesse. C'erano regioni caratterizzate da nuclei, che sostenevano la stabilità nella regione. Tutto grazia a una grande mobilità.
Espansione è stata permesse anche grazie all'utilizzo di indumenti, testimoniato dal ritrovamento di antichi aghi per cucire, e costruzione di rifugi artificiali.
Il primo sito con un reperto che mostra una continuità genetica con l'essere umano attuale europeo è il sito di Kostienki, in Russia datato a 39-37 Ka. Kostienki 14 presenta una continuità genetica con popolazioni successive, come quelle rinvenute a Dolni Vestonice, in Repubblica Ceca, datato a (27-26 Ka).
Tuttavia, l'ultimo massimo glaciale (25-19Ka) affievolisce questa continuità genetica. Le popolazioni si frammentano e si rifugiano a latitudini più meridionali. Appr.
Non è chiaro il contributo genetico di queste popolazioni agli europei post LGM. Il segnale pare rimanere nelle culture caratterizzate da industria sultreana, che abitano la penisola iberica durante il LGM, e le culture successive caratterizzate da industria maddaleniana che poi si sposteranno verso Nord-Est.
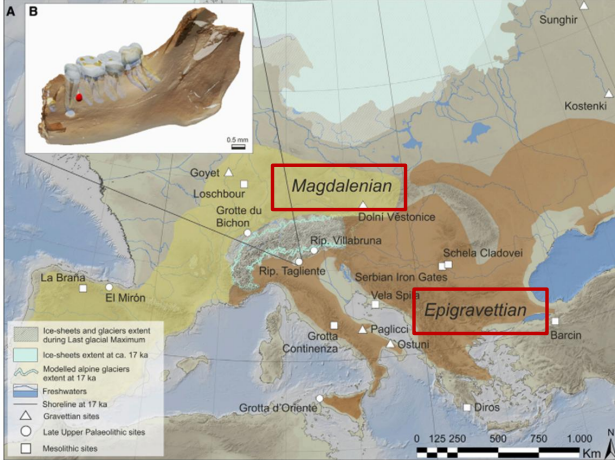
Approffittando delle regressioni marine, l'umanità raggiunse l'America settentrionale. I momenti favorevoli per questo passaggio devono essere stati negli intervalli tra i 70-35 Ka e i 26-9 Ka.
Precedenti ipotesi non prevedevano la possibilità di espansione prima dell'artefatto datato più anticamente, le punte di Clovis datate a 13-11 Ka, Appr.
Ipotesi attuali invece ipotizzano come prima presenza i reperti di Bluefish cave, datati a 24 Ka. Tuttavia questi primi sapiens amerindi devono essersi stanziati per circa 10 Ka vicino allo stretto di Bering, per poi espandersi rapidamente attorno a 15 Ka. Numerosi altri siti sostengono l'ipotesi di una colonizzazione Pre-Clovis.
9 Quante specie di Homo conosci: distribuzione spaziale e cronologica
| Più antico | Più recente | Areale | |
|---|---|---|---|
| H. habilis | 2,3 Ma | 1,8 - 1,5 Ma | Africa SudOrientale |
| H. rudolfensis | 2,5 Ma ° | 1,6 Ma ° | Africa Orientale |
| H. erectus s.l. | 2 Ma | ergaster 1 Ma s.s 117-108 Ka |
Africa, poi Asia e Europa in alcune sue forme |
| H. antecessor | 850-800 Ka | / | Sierra de Atapuerca |
| H. heidelbergensis (1200cc) | 700 Ka | 200 Ka | Africa, espanso in Eurasia |
| H. naledi | 335-226Ka | °°° | SudAfrica |
| H. sapiens (1450cc) | 315 Ka °° 233 Ka |
:D | Evolutosi in Africa, espanso in tutto il globo |
| H. neanderthalensis | 250 Ka | 40 Ka | da Europa, espanso in Vicino Oriente e Asia Centrale |
| Denisoviani | 250 Ka | 50 Ka | Asia, dai Monti Altai al sudEst. |
| H. floresiensis | 1 Ma | 90-50 Ka | SudEst Asiatico, Isola di Flores |
| H. luzionensis | 67 Ka | °°° | Filippine |
° Molto dibattuto
°° Se si considerano i reperti di Jebel Hiroud propriamente come H. sapiens
°°° Un solo reperto / sito
H. habilis
Appr. / slide
Il reperto più antico attribuito a H. habilis è datato ad 2,3 Ma, in Etiopia. a 2 Ma è datato il cranio di Koobi Fora, in Kenya. Il più recente è stato rinvenuto in SudAfrica ed è datato tra i 1,8 e 1,5 Ma.
Spazialmente, grazie ai reperti incontrati, era distribuito in Africa SudOrientale. Potrebbe essere originato in Africa Orientale (Etiopia, Kenya) ed essersi spostato in SudAfrica in un secondo momento.
H. rudolfensis
Appr. / slide
C'è molto dibattito sulla specie. Sostanzialmente i reperti non sono numerosi e sono di difficile attribuzione, manca completamente lo scheletro postcraniale.
I reperti possibilmente attribuiti fanno pensare a un range compreso tra i 2,5 Ma e i 1,6 Ma
H. erectus s.l.
Appr. / slide / slide
I reperti più antichi sono adatati attorno i 2 Ma, con un cranio parziale ritrovato a Drimolen, in SudAfrica, attribuito alla sua forma africana, H. ergaster.
Importanti reperti sono il Ragazzo del Turkwana e le Impronte di Ileret, entrambi datati attorno ai 1,5 Ma.
L'areale di H. ergaster è molto vasto, e raggiunge l'Africa Settentrionale. A questa specie è attribuito il primo evento di dispersione al di fuori dell'Africa.
H. antecessor
Appr.
Un unico sito fornisce reperti attribuibili alla specie, si trova alla Gran Dolina, Sima de Atapuerca, ed è datato tra i 850 e 800 Ka.
H. heidelbergensis
Appr. / slide
In seguito a una lacuna del record fossile, tra i 900 e 600 Ka, emerge H. heidelbergensis. Siamo nel Pleistocene medio.
I primi reperti attribuiti alla specie si incontrano in Africa, a Bodo, in Etiopia, datati a 600 Ka. E' ben distinguibile per l'elevata capacità cranica (1200cc). I reperti datati più recentemente, potrebbero attribuire la durata della specie nel continente africano fino ai 300 Ka, avrebbe così coesistito con H. sapiens e H. naledi.
.
H. heidelbergensis deve essere stato un taxon molto variabile morfologicamente e di ampia distribuzione geografica e deve essere vissuto per tutto il Pleistocene medio, tra i 600Ka e i 200Ka.
In Europa i primi reperti spaziano tra i 700 Ka (non è diretta l'attribuzione), mentre i primi resti spaziano tra i 480 Ka e i 250 Ka. In Asia ci sono molti reperti, ma nessuno è direttamente attribuito a H. heidelbergensis.
Non è chiarissimo da quale specie sia evoluto H. heidelbergensis, ma l'ipotesi più probabile parla di un origine da H. ergaster.
H. neanderthalensis
Appr. / slide / slide
I Neanderthal sono distinguibili in "N. arcaici" ascritti al Pleistocene medio, e "N. classici" concentrati nel Pleistocene superiore, a seconda del mosaico di morfologie che mostrano.
Il record fossile è molto ampio e fornisce una varietà morfologica molto vasta.
Cronologicamente hanno popolato l'Europa per un lungo tempo prima dell'arrivo di H. sapiens, e si sono diffusi in Vicino Oriente e in Asia Centrale.
Hanno una morfologia particolare, ben distinguibile da H. sapiens. Appr.
Denisoviani
Appr. / slide
I denisoviani non hanno ancora un olotipo, perciò non sono ancora classificabili come specie. Tuttavia indagini sopratutto di tipo genetico mostrano l'esistenza di una popolazione ben distinta dai coevi H. neanderthalensis e H. sapiens.
I Denisoviani sono un fenomeno unicamente asiatico. La divergenza coi Neanderthal/Sapiens deve essere avvenuta tra i 473-443 Ka.
La maggior parte dei resti attribuiti ai denisoviani viene dalla Grotta di Denisova, con datatazioni molto varie (Dai 250 ai 50 Ka).
Nel SudEst asiatico inoltre ci sono molti altri reperti che potrebbero essere attribuiti ai denisoviani, come il cranio di Cranio di Harbin, la Mandibola di Xiahe e la Mandibola di Penghu, anche il dibattito è ancora attivo e molti resti asiatici sono sotto revisione.
Ciò però indicherebbe una notevole diffusione dei denisoviani nel continente asiatico, anche per un lasso di tempo molto lungo.
H. sapiens
Appr.
Il sito più antico è quello di Jebel Hiroud, in Marocco, datato attorno ai 315 Ka, dove son stati trovati reperti che mostrano una morfologia mandibolare e dentale moderna, seppur rimane arcaica la forma di cranio e neurocranio (è assente la globularità del cranio). Il fossile più recente con cranio moderno è datato a 233 Ka, nella valle del fiume Omo, in Etiopia. C'è da considerare che l'origine di H. sapiens è considerato come un evento panafricano, quindi non circoscritto in un'area ristretta. A Herto Bouri (160-150 Ka) viene trovato il primo cranio moderno completo, che mostra splancnocranio e neurocranio.
Ad Homo sapiens è associato inoltre il terzo evento Out of Africa. L'areale, direi, che lo conosciamo.
H. floresiensis
H. naledi
Specie molto peculiare siccome, nonostante gli unici reperti la collochino tra i 335-226 Ka in SudAfrica, aveva caratteristiche molto primitive, assimilabili ai primi australopitechi, come notevoli capacità di arrampicamento a scapido del bipedismo, piccole dimensioni cerebrali (seppur una possibile anatomia del cervello simile ai contemporanei). Non è chiaro se fosse una specie diffusa al di fuori del sito di ritrovamento. Potrebbe essere evoluta da H. erectus
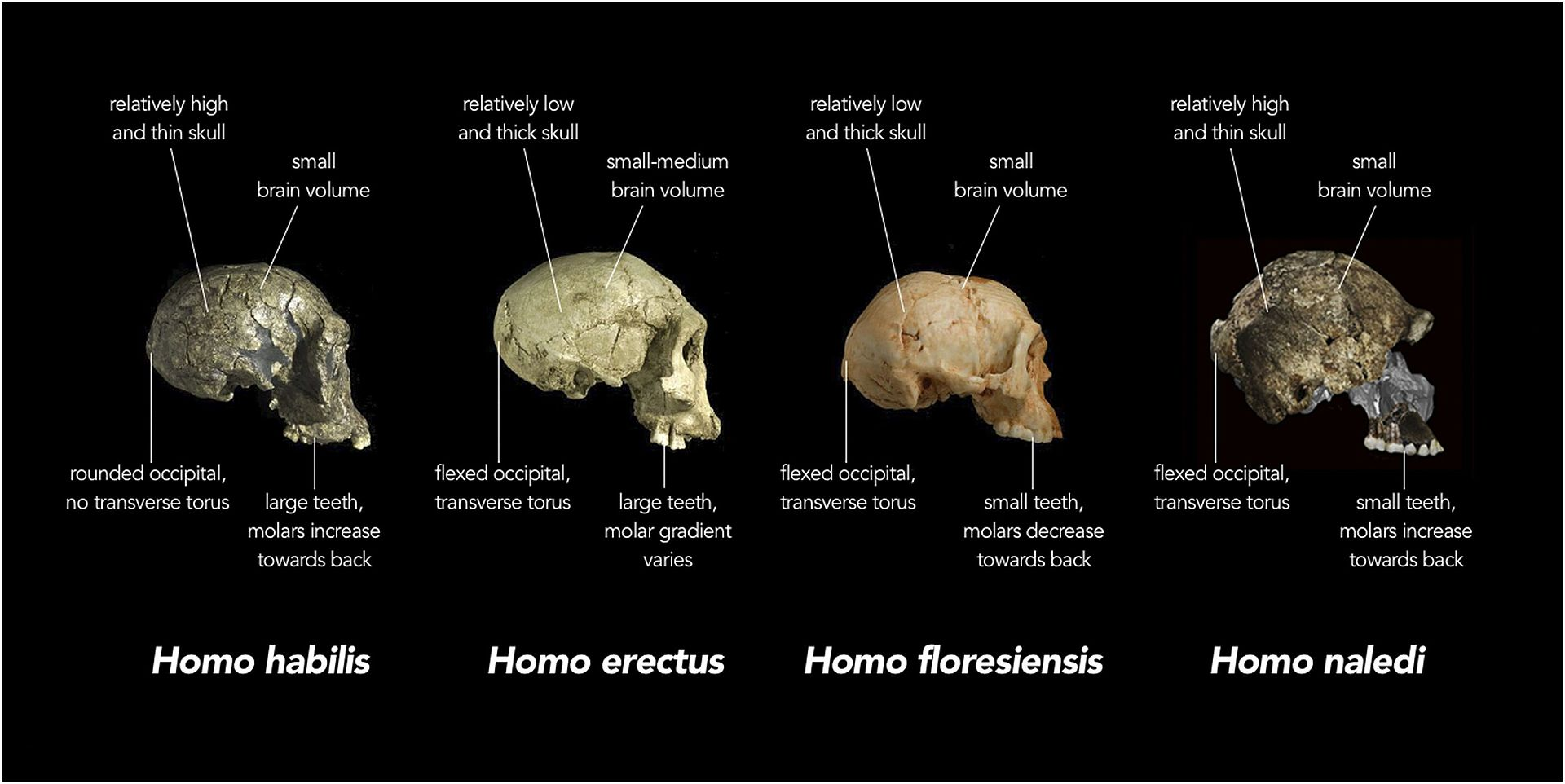
Wikipedia
H. luzoniensis
Callao Cave, datato a 67 Ka, quindi la prima colonizzazione attestata delle filippine.
Queste isole possono essere incluse o escluse nelle faune delineate dalla Linea di Wallace se si prende in considerazione le modificazioni di Huxley
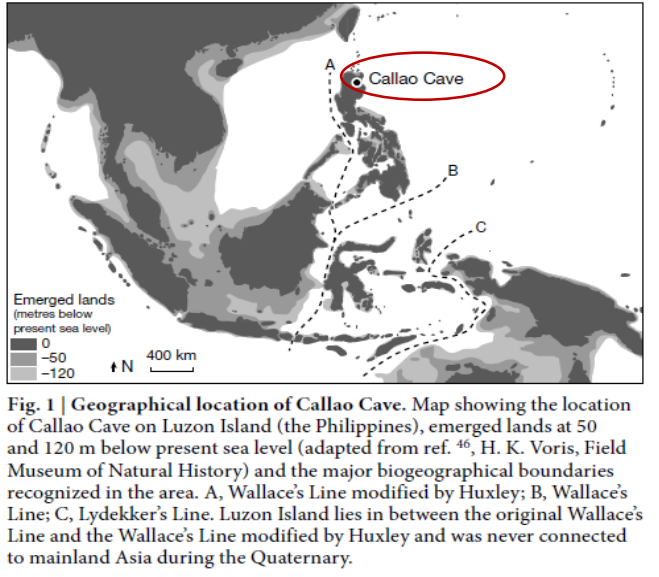
Sono pochi frammenti, con caratteristiche di nanismo insulare. Non rientrano nella variabilità già note di altre specie. Non è chiaro da chi derivi. Comunque indica un popolamento di queste regioni tra Pleistocene superiore e medio.
10 I Neanderthaliani
Qui linkerò semplicemente i miei appunti, vedrò se sintetizzare
Originato da Homo heidelbergensis circa 250ka, (1400cc). La cesura tra viene messa attorno ai 250'000, quando sono visibili tutti i caratteri tipici neadertaliani:
- Encefalizzazione estrema, scatola cranica voluminosa e allungata
- Protuberanza dell'occipitale
- Marcato toro sopraorbitario
- Fronte sfuggente
- altre caratteristiche apomorfe del Neanderthal.
La morfologia del cranio è ancora molto arcaica, allungata e poco globulare. Hanno caratteristiche dello scheletro facciale tipiche dei Neanderthaliani.
Sono un fenomeno Europeo con dispersione in vicino oriente e Asia centrale in fasi più tardive.
Il record fossile e manifestazioni culturale è molto ricco.
Si estingue circa 40'000 anni fa, non senza lasciare la loro traccia nel DNA umano.
I dati che hanno permesso una ricostruzione filogenetica tra Neanderthaliani e Homo sapiens, stimano una divergenza avvenuta tra i 750'000 e 550'000 anni fa.
Sono stati evidenziati inoltre distinti eventi di ripopolamento e dispersione, probabilmente causato da estinzioni locali e successive dispersioni.
Caratteristiche morfologiche
Sicuramente a queste particolari morfologie trovano motivi in processi di deriva genetica e inibridazione. Si ipotizza anche la possibilità di processi ontogenetici diversi. Però lo svezzamento pare essere simile a quello dell'uomo moderno (attorno ai 5-6 mesi). In Neanderthal non si riscontra un vero e proprio processo di invecchiamento, con la osteoporosi classica di H. sapiens. Tuttavia è addicibile anche a processi culturali (il mantenimento degli anziani)
Craniali
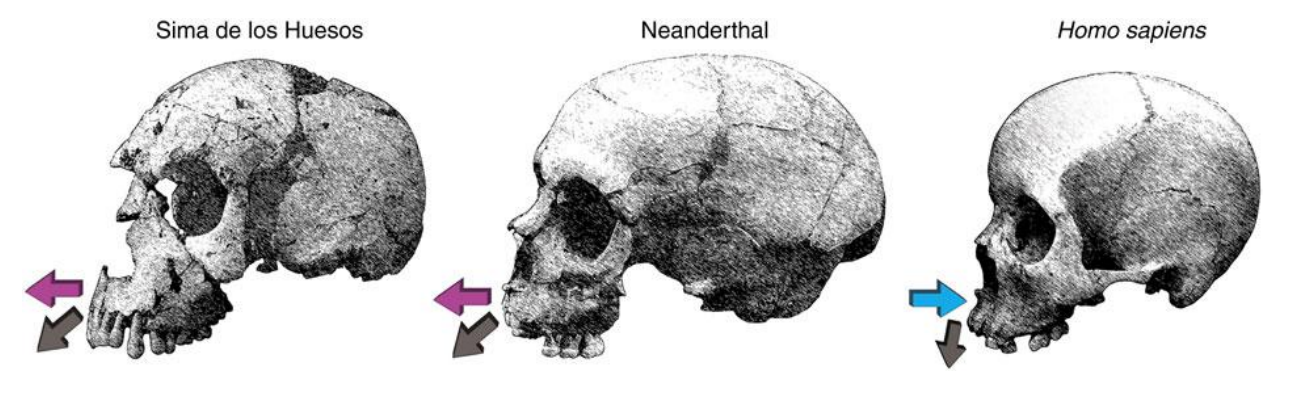
Capacità cranica maggiore di H. sapiens (1500-1600). Il cranio è allungato, rispetto a quello di H. sapiens. La faccia, soprattutto la regione mediofacciale è "spinta in avanti". I neanderthaliani sono facilmente riconoscibili.
La regione sopraciliare e sopraorbitale del torus sono fuse, per questo è detto torus continuo. Per H. sapiens si parla di rilievo, non di torus, siccome è solo sopraorbitale.
Le orbite sono tonde, c'è prognatismo medio-facciale (Cioè la parte zigomatica e mediofacciale è spinta in avanti) Ciò causa l'appiattimento dell'osso mascellare.
Il mascellare è in estensione. Mentre la fossa canina è completamente assente.
In
Neanderthal è assente il mento. Il foro mentoniero in Neanderthal è sotto il primo molare (M1), in sapiens è in (P2-P1)
Sicuramente il morso dei neanderthal è distinto, a "tenaglia". I denti hanno una superficie molto ampia e uno spessore importante. Le radici sono molto fuse, indicando una forte pressione all'utilizzo dei denti come ulteriore "mano".
Nella parte posteriore del cranio, è molto caratteristica la fossa soprainiaca
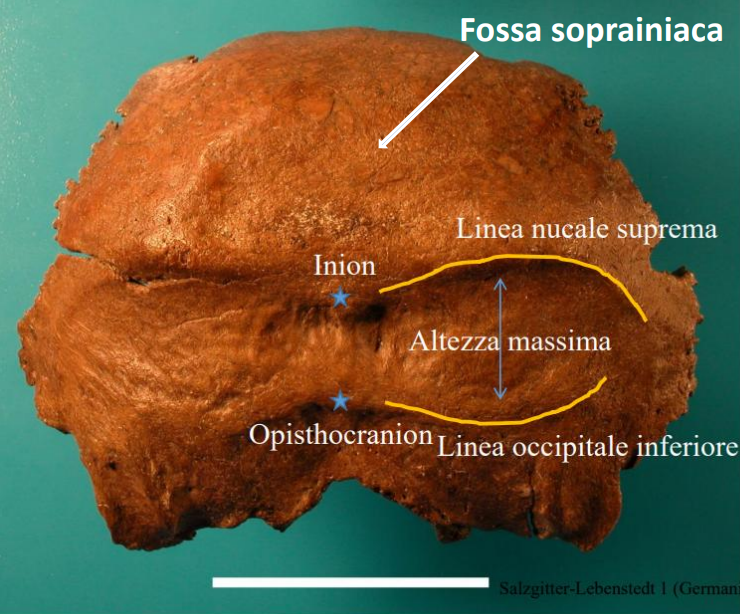
Postcraniali
Diagnostici:
Il ramo pubico è più lungo e sottile che in sapiens. L'inserzione del muscolo piccolo rotondo avviene su una faccia distinta della scapola rispetto a Sapiens. La sezione laterale del femore è rotonda e non triangolare come in Sapiens
Tronco e bacino più largo.
Le ossa sono molto robuste, cioè ispessite, se comparate a quelle di sapiens.
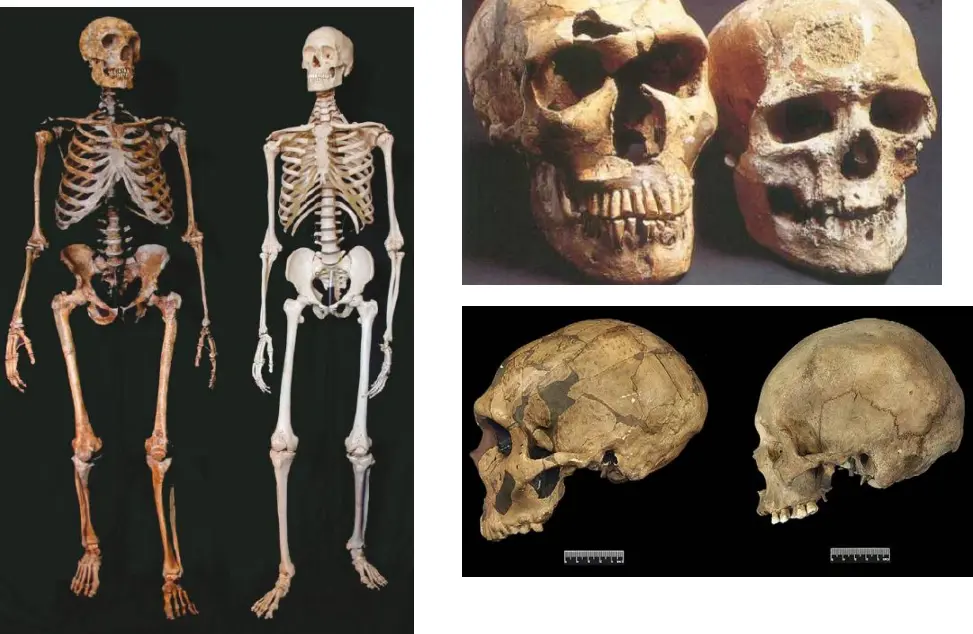
Postura e locomozione
Grande gluteo
La sua contrazione estende l'arto inferiore. In H. sapiens è sulla faccia posteriore del femore.
Nei Neanderthaliani questo muscolo è inserito più lateralmente e l'inserzione nel femore ha forma più lanceolata. Può essere causato da una diversa inserzione del muscolo e diversa posizione del tendine. Data dall'allungamento del pube e allargamento del bacino. Il muscolo e il tendine sono in frazione.
Dai reperti di Krapina, un muscolo così importante ma così distinto nel suo attacco, implicherebbe un distinto sforzo meccanico. Nei Neanderthal è in trazione, con una superficie verosimilmente coperta di cartilagine.
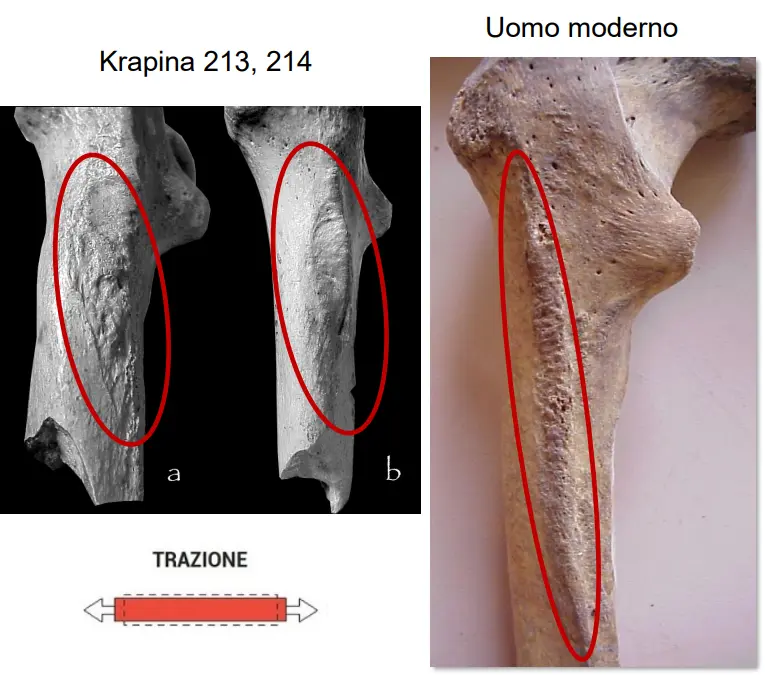
Ciò implica una meccanica distinta, un funzionamento di questo organo molto diversa. Ha un sedere piatto, molto stirato lateralmente. Può comportare che in Neanderthal non camminassero eretti come no, ma più ????
Muscolo vasto intermedio
E' visibile l'impronta sui femori adulti. Sulla faccia interiore del femore è presente un muscolo estensore del ginocchio. Potrebbe implicare una diversa distensione del ginocchio
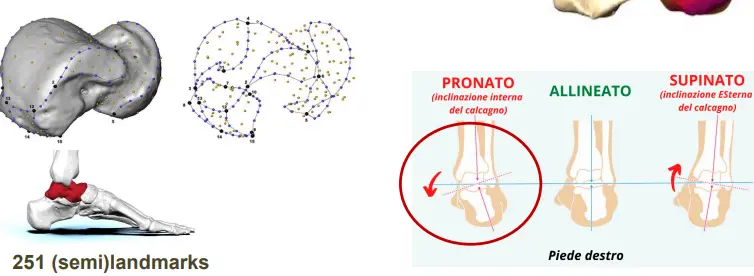
Piede
Un osso di cerniera, nella sua faccia di articolazione col calcagno ha una distinta attaccatura e diversa densità del tessuto osseo. Potrebbe essere legato a una maggiore pronazione del piede, con un piede più piatto.
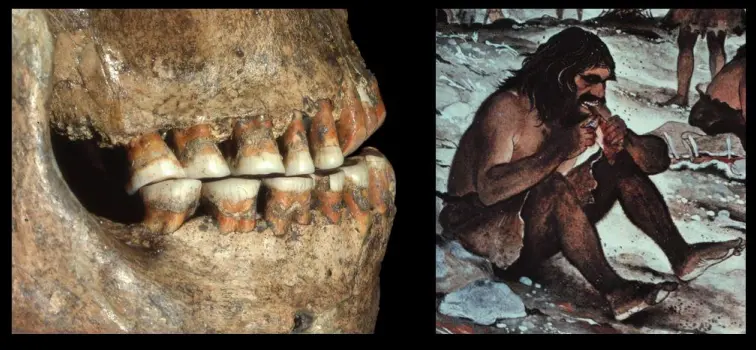
Comportamenti alimentari
Denti molto usurati, probabilmente perchè venivano utilizzati anche per attività extra alimentari. Dal sito di Shanidar.
Sempre a Shanidar è stato trovato un corpo a cui era stato amputato un braccio, e probabilmente cieco (caratteristiche degli orbitali). Testimonia cura e cooperazione da parte del gruppo sociale.
Piccole fratture dello smalto o asportazioni della corona. Dovute in parte alle attività masticatorie. Molto probabilmente dati dall'uso dei denti come "terza mano", o in generale attività extra alimentatari.
Ci sono studi sul tartaro dei denti Neanderthaliani. Si accumula al margine della gengiva, in prossimità delle ghiandole salivari.
A El Sidròn si riesce a riscontrare una alimentazione vegetariana. Dallo studio del microbioma si è riuscito a scoprire l'utilizzo di una specie di aspirina, per la cura di ascessi.. A Spy cave in Belgio una maggiormente onnivora.
Linguaggio articolato
Studi recenti attestano capacità di linguaggio e auditorie simili a quello di H. sapiens. Ricostruendo la struttura del canale, dell'orecchio medio e orecchio interno, gli studiosi sono riusciti a trasferire suoni su questi modelli virtuali, dimostrando capacità molto simili a quelli di H. sapiens. Di contro, gli ominidi di Sima de los Huesos (più arcaici) non avevo capacità di linguaggio simili.
La vocalizzazione d'altra parte era un carattere sicuramente già plesiomorfico
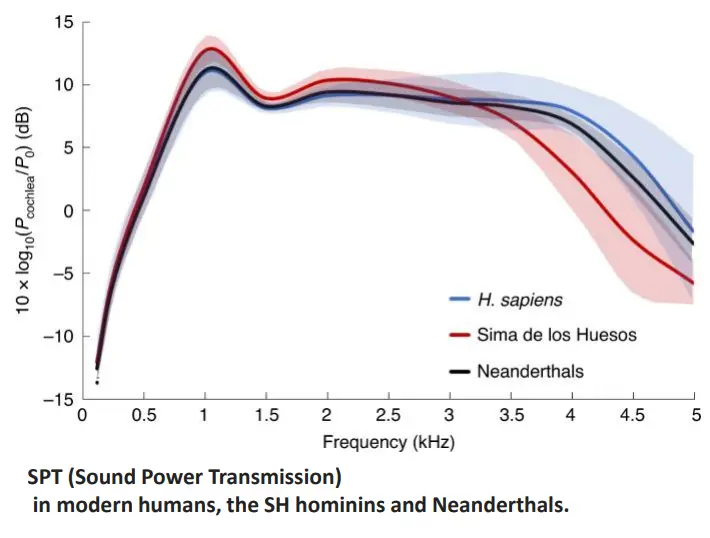
Il reperto Kebara 2 mostra la morfologia dell'osso ioide identica a H. sapiens, che indica una mobilità molto simile della lingua a H. sapiens.
Mobilità in Asia
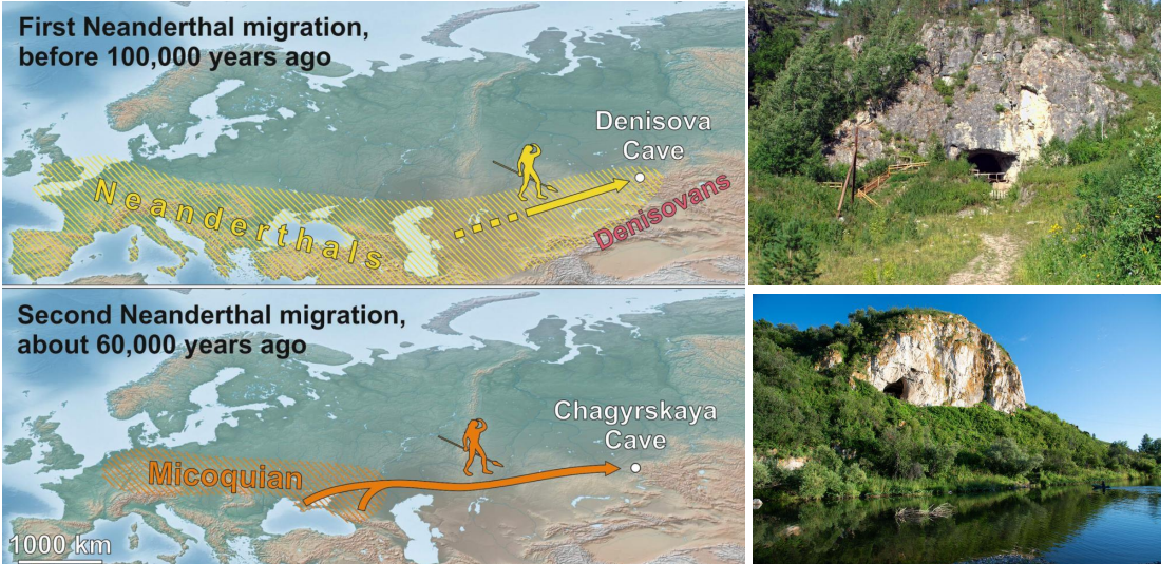
E' nota la presenza e la mobilità di neandertaliani in Asia. Oltre alla grotta di Denisova c'è anche la grotta di Chagyrskaya. Una prima dispersione è attestata a 100'000 anni fa, mentre l'altra ai 60-50'000 anni fa. Si son portati una industria Micoquiana,
tipica del paleolitico medio, rinvenuta anche in Crimea.
Convivenza con sapiens
Comparsa della specie
Già a Sima de los Huesos (430 Ka) erano visibili i primi accumuli di caratteri Neanderthaliani. (Coxa vara e Fossa oleocranica larga)
Anche a Saccopastore 250'000.
Un po' in tutta Europa tendono ad accumularsi nel tempo caratteristiche Neanderthaliane.
Questi processi sono stati velocizzati da i processi glaciali. Il susseguirsi di cicli climatici frammentavano e riducevano le popolazioni, causando processi di deriva genetica, che fissavano certi caratteri.
La specie ancestrale (H. heidelbergensis) sarebbe stata sottoposta a fasi di isolamento geografico con ricorrenti crisi demografiche, connesse all'alternanza di fasi glaciali e interglaciali. Ad ogni fase glaciale solo una piccola frazione della diversità sopravviveva, fissando queste varianti. Al ritorno di condizioni favorevoli queste morfologie si riespandevano, per poi tornare a contrarsi. Così si sono potute accumulare caratteristiche in senso Neanderthaliano.

Ipotesi anagenetica
Si ipotizza possa essere avvenuto un evento anagenetico. Cioè che siano comparse tante caratteristiche, poi accumulate nel tempo e che ha portato al Neanderthal.
Ipotesi organismica
Si ipotizza anche un modello organismico. Proposto da Antonio Rosas, propone che ci siano state poche mutazioni che hanno portato alla comparsa di alcuni fenotipi, per cui però data l'interdipendenza delle strutture del cranio, sono comparse assieme. Quindi una cascata di caratteristiche che deriva da un numero limitato di eventi di mutazione.
Reperti
Arcaici
Sono distinti in N. arcaici (da circa 250Ka) e classici da (120-49Ka)
Abitavano steppe e foreste.
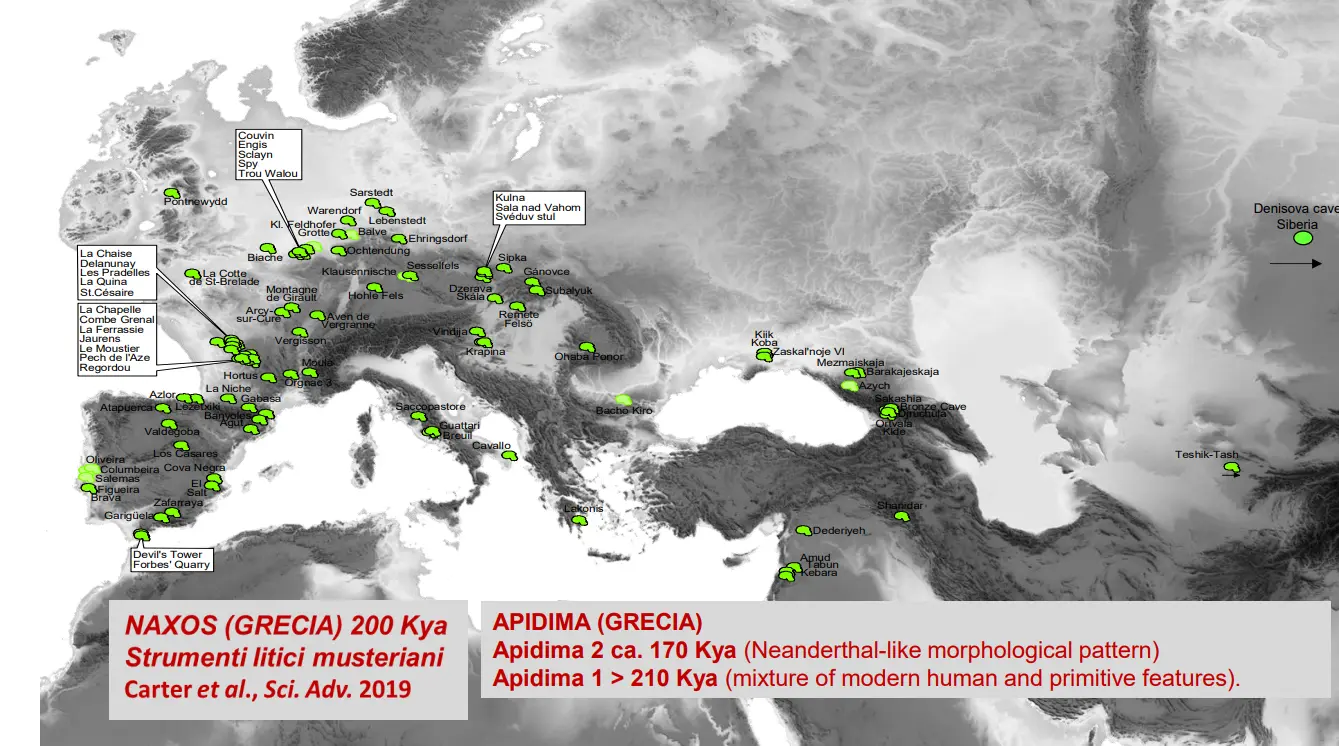
Dal punto di vista delle datazioni, ci sono resti piuttosto antichi. C'è un cranio parziale non direttamente attribuito a Neanderthal ma che ne porta molte caratteristiche, trovato in vicino oriente, e datato tra i 500 - 200 Ka. Potenzialmente quindi potrebbe essere il resto più antico, ma permangono molti dubbi.
Datati più recentemente sono i crani trovati a Saccopastore con datazioni attestati ai 250Ka.
Nelle grotta di Denisova sono stati trovati reperti dai 200 ai 280'000 anni fa.
I cosiddetti "ultimi neanderthal" attestati ai 40Ka, in Europa Orientale. In portogallo è stato trovato un reperto con molti caratteri ibridi, attestato a circa 24'000 anni fa. I gruppi dell'Europa orientale hanno rappresentato un coul de sac (chevuoldire??)
Medio Oriente
Tra i 400-200Ka in Palestina ci sono fossili con caratteristiche arcaiche, simili ad altre popolazioni arcaiche ed altre con caratteri particolari. Sono tutti reperti non facilmente attribuili a H. neanderthalensis o H. sapiens. Sono zone di sosta e zone dove si stabiliscono a lungo termine distinte popolazione. Qui l'H. sapiens dopo l'uscita dall'Africa incontra i Neanderthaliani già presenti, arrivati dall'Europa.
Qui popolazioni hanno convissuto e condiviso riti funerari e tecnologie litiche
- Qesem Cave, 400-200'000.
- Nesher Ramsla, 140-120'000.
- El Zuttiyeh 250ka (Neanderthal)
- Misliya Cave, 194-177 Ka. (H. sapiens)
Krapina
Krapina, sito molto ricco, 930 resti ossei e dentari, in Croazia, attribuito a 130Ka.
Non sono stati trovati però ossa o individui completi, solo fratture e tagli. Può essere dovuto a come è stato scavato il sito (dal 1899 al 1905). Tutti i resti sono fossilizzati. C'è scritto Homo sapiens perchè inizialmente i neanderthal fossero una sottospecie. Molto resti litici e faunistici.
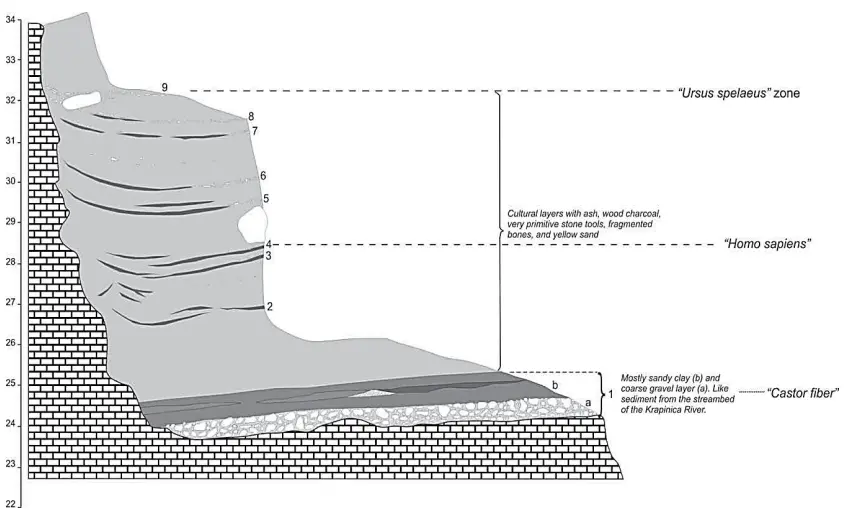

E' ricca di reperti litici e ossei che testimoniano attività. Ci sono tagli che testimoniano attività di scuoiamento e disarticolazione.
Ossa con blunt trauma, le schegge son rimaste attaccate, quindi indica che sia stato fatto su osso fresco, probabilmente per conservare il midollo
!!!

El Sidròn
49'000 dove son stati ritrovati 2500 resti ossei e dentari, appartenente a fasce distinte. Pochi resti litici. Grotta nelle Asturie spagnola. Era probabilmente un luogo funeriario. E' stato possibile estrarne il DNA, dimostrando che era un gruppo familiare.
Saccopastore
A Saccopastore sono stati rinvenuti due crani datati a 250 Ka.
Ha caratteristiche tipiche dei Neanderthal
Altamura
Ciccillo Lobaccaro
Apidima
In Grecia.
Due reperti, uno di H. sapiens e uno attribuito a H. neanderthal. Però è più recente quello neanderthaliano. Può supporre alternanze di Neanderthal e Sapiens, così come una fuoriuscita dall'Africa di Sapiens antecedente a quanto si pensava precedentemente.
- Apidima 1
- Globulare, tipico sapiens
- Apidima 2
- Torus importante, profilo circolare diagnostico dei Neanderthaliani. L'occipitale è curvo.
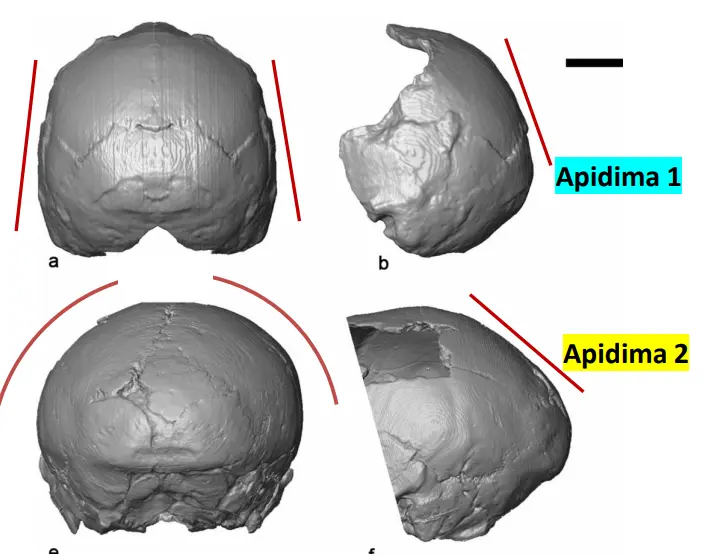
- Torus importante, profilo circolare diagnostico dei Neanderthaliani. L'occipitale è curvo.
Classici
Grotte Mandrin
Come si vede nella mappa, gli ultimi Neanderthaliani erano collocati in Europa occidentale.
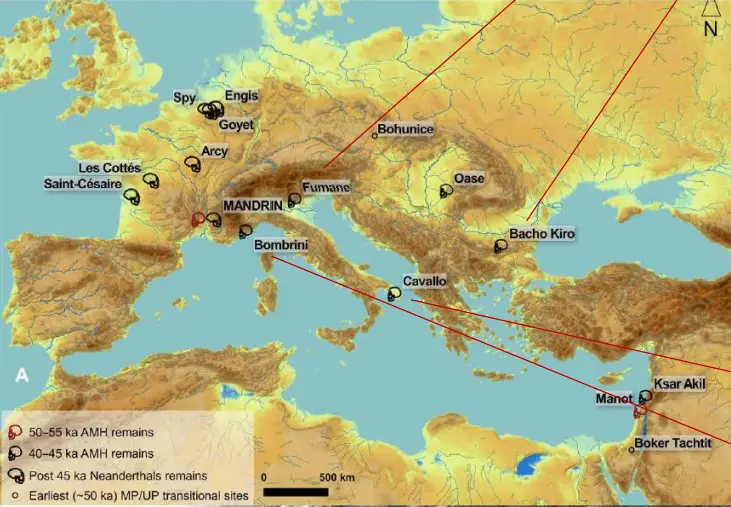
Nella Grotta Mandrin sono stati trovati crani di H. sapiens datati a 56-51'000 anni fa, mentre successivamente son stati rinvenuti Neanderthal, datati post 45ka.
Eccetto il caso di Apidina, indicherebbe un ingresso di sapiens in Europa più antico di quanto noto.
Si rinvengono manufatti litici non conosciuti al di fuori dell'Africa in quel tempo.

Aumenta quindi il tempo di convivenza tra questi due ominidi (a circa 10'000 anni). Tutte le industrie di questo periodo diventano critiche dal punto di vista dell'artefice, di chi lo ha realizzato. La Tecnica Levallois è già diffusa e le tecnologie sono simili.
Spostamento in Asia
Starosele
In Crimea son stati rinvenuti reperti, datati attorno ai 46-45 Ka.
con la spettrometria di massa si riescono a distinguere frammenti di artefatti umani da resti animali. Una buona percentuale dei frammenti aveva ancora collagene.
Denisova e Chagyrskaya
Sono stati trovati reperti datati rispettivamente a 200-100 Ka e 60-50 Ka, perciò si può supporre che siano avvenute più ondate di espansione dall'Europa verso l'Asia centrale. A corroborare questa teoria ci sono evidenze di diffusione dell'industria micoquiana (bifacciali senza tecnica Levallois) nella zona tra Crimea e monti Altai.
Cultura e comportamenti simbolici
Segni paralleli tra loro in un teschio di una donna, rinvenuta a Krapina.
Sepoltura
Quando i Neanderthaliani seppellivano, ancora non erano presenti sepolture di H. sapiens, perchè sono attestate successivamente (fuori dall'Africa).
Le sepolture comunque rispecchiano dal punto di vista culturale un preadattamento a una certa forma di insediamento, in quanto si gravita attorno un luogo. Rappresenta un legame al territorio, una transizione verso il Neolitico
Grotta di Tabun,
attestazione della sepoltura più antica (170-120'000 anni fa) nota. Attribuita a Neanderthal. Questo per la posizione degli arti inferiori, flessi. C'è una serie di caratteristiche che indicano una possibile sepoltura. L'intenzionalità si rinviene anche dalla scavatura della fossa. Non c'è però corredo, che indicherebbe attività funeraria vera e propria.
Grotta di Qafzeh
Sepoltura più antica di H. sapiens. Si mostra anche una importante intenzionalità, vengono sepolti anche due individui (donna e bambino) assieme. Potrebbe indicare una mortalità congiunta, o che uno sia stato sacrificato per accompagnare l'altro come corredo.
Tecnologia litica
Associato ai Neanderthal si parla di Musteriano (Modo 3), perfezionano la tecnica Levallois.
Ci sono testimonianze dell'utilizzo della tecnologia litica per l'accensione del fuoco.
In Toscana si attestano anche strumenti in legno induriti tramite il fuoco.
Altre espressioni culturali
- Le grotte comunque attestano una certa organizzazioni e costruzioni, utili all'organizzazione del luogo.
- Utilizzo del fuoco nella quotidianità
- Sono attestati anche possibili ornamenti.
- Primi a realizzare strumenti specializzati in osso
- ttestata l'utilizzo della colla con catrame di betulla (50'000)
- Utilizzo di pigmenti scuri, utilizzati probabilmente per dipingersi e tatuarsi.
- Pitture in grotta (probabilmente associati a sapiens)
- In gibilterra, in una grotta è stato trovato un pattern che dimostra una certa regolarità
Nel periodo compreso tra i 50'000 e 40'000 anni fa, contestualmente all'ingresso di Homo sapiens in Europa, ci sono reperti possibilmente attribuibili a H. neanderthal. E' però difficile attribuire i reperti, siccome diversi gruppi di Neanderthal avevano distinte tecnologie litiche.
Ai Neanderthal è stata anche attribuita la produzione di artefatti associati all'industria Castelperroniana.
Inoltre è frequente l'utilizzo di artigli d'acquila, frequenti in numerosi siti.
Fumane
Luogo di transizione, dove son convissuti Neanderthal e Sapiens.
Son state trovato ossa di rapace, con una serie di tagli e lesioni, che più che per uso alimentare, potrebbero indicare l'asportazione di penne, magari usate per ornamento.
Paleogenetica
Il genoma dei neanderthal dei monti Altai, ha un 6% di tracce di genoma umano, ma una linea specifica di Sapiens usciti dall'Africa verso l'Eurasia prima di 100'000 anni fa, ma poi scomparsa. (Vedi anche Monti Altai)
Ibridazione tra Neanderthal e Sapiens
Grazie agli studi di Svante Paabo sappiamo che una piccola percentuale del DNA umano deriva dall'incrocio con distinte popolazioni. Noi definiamo sapiens e neanderthalensis come specie distinte, e si ritengono tali. Hanno ancestralità comune con heidelbergensis, ma i 300'000 anni che intercorsero tra l'origine dei due cladi non è stata sufficiente da impedire l'ibridazione.
Sono stati gli studi di paleogenomica a dare informazioni sull'ibridazione di Neanderthal. Hanno permesso anche di allargare il range di distribuzione di Neanderthal verso l'Asia.
Ora si attesta che tra l'1% e il 4% del genoma umano sarebbe di origine neandertaliana.
L'ibridazione è quindi successa tra i 60 e 50ka, dopo la fuoriuscita dall'Africa e prima della diversificazione delle popolazioni di Sapiens.
Non pare ci sia stata una ibridazione con alcune popolazioni subsahariane, forse a indicare una espansione unidirezionale verso l'Asia e l'Europa. Tuttavia col ritorno di Homo sapiens sotto il Sahara sono state portate anche le tracce del DNA neanderthaliano.
Per alcuni motivi, è stato perduto il DNA mitocondriale, è chiaro che ci fossero certi ibridi favoriti: maschi N. e femmine di H.s, se nella prole nascevano maschi, erano meno fertili delle femmine.
Non c'è traccia del cromosoma Y nei N nei maschi di H.s, per una mutazione. Il DNA di Neanderthal presenta mutazioni in tre diversi geni immunitari, che potrebbe aver causato risposte immunitarie delle donne H.s incinta, causando aborti dei figli maschi, che se nascevano probabilmente non erano sani.
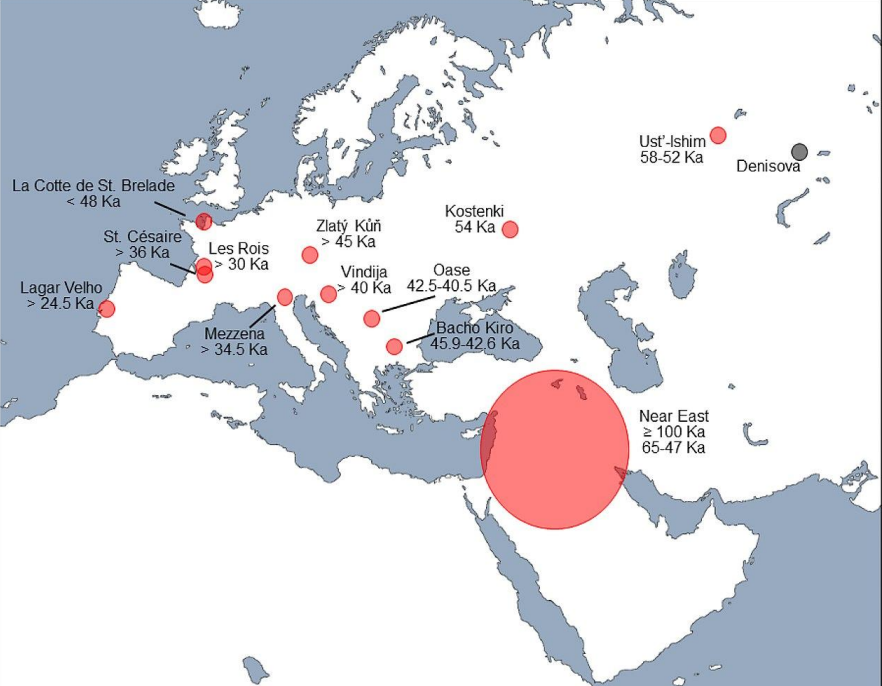
Gli incroci hanno anche mostrato che l'ancestralità neandertaliana asiatica è presente in percentuale maggiore in Asia. Però si vede che invece in Europa l'ibridazione è più recente. Questo perchè nuovi ingressi di H. sapiens in Europa hanno "diluito" la percentuale di DNA Neanderthaliano in Europa.
Struttura sociale
Popolazioni isolate di piccoli gruppi, chiaramente endogamiche. 13 Neanderthal ritrovati in due grotte, nelle montagne Altai nella Siberia del Sud. Avevano tecnologia micoquiana.
Erano componenti di una stessa famiglia, si è riuscito a capire come fosse composta.
Si mostra anche l'eteroplasmia dei mitocondri, condivisa dagli individui, ad indicare parentela tra questi.
Rischi
Una percentuale di DNA neanderthaliano coincidono con un maggior rischio di ospedalizzazione dal Covid19. Allo stesso tempo altre parti del genoma sono associati ad una protezione da Covid19.
11 I Denisoviani
Sono un fenomeno esclusivamente asiatico.
Alcuni reperti trovati in Cina potrebbero appartenere a questa specie.
Potrebbe derivare da H. heiderbengensis o da H. erectus, non è ancora completamente chiaro.
Come H. neanderthalensis, si estingue 50'000 anni fa, non senza lasciare la loro traccia nel DNA umano, in seguito all'ibridazione con H. sapiens che andava espandendosi nei supo terrotpro.
Si stima che la divergenza con i neanderthal sia avvenuta circa 445-473 Ka.
Non è stato ancora trovato l'holotipo, per questo non è ancora descritto a livello di specie
Non è ancora stata creata un'associazione tra morfologia (esistono pochi reperti) e caratterizzazione genetica (è stata possibile su una quantità molto ristretta di resti)
In generale il contesto asiatico è sotto grande revisione.
Grotta di Denisova
Denisova cave
Nei monti Altai, qui si vede che Homo neanderthalensis e denisoviani si sono potuti incrociare.
L'intero genoma mitocondriale Denisoviano è stato sequenziato nel 2010-2012, Nel 2014 l'interno genoma neandertaliano, da un reperto nei monti Altai.
Molti reperti frammentari, gli studi sono cominciati dal 2010. L'appartenenza alla specie di questi frammenti è stata possibile grazie ad indagini di tipo biomolecolare. Si è visto che l'Asia era abitata da una popolazione arcaica che ha lasciato la propria eredità genetica nel DNA neandertaliano.
La grotta è stata abitata per molto tempo, infatti i reperti hanno datazioni anche molto diverse:
- Denisova 3, 4 - 100-54 Ka circa
- Denisova 2 - 150 Ka circa
- Denisova 8 - > 100 Ka
- Denisova 19, 20, 21 - 250 Ka
I reperti Denisova 4 e Denisova 8, che sono due molari, hanno denti di dimensioni maggiori dei Neanderthaliani.
I Neanderthaliani di questa grotta hanno lasciato pochi frammenti.
Un aspetto importante è che il 6% del genoma dei neanderthaliani dei monti altai (datati 140-120'000) contengono un'introgressione di DNA sapiens di gruppi usciti dall'Africa prima di quanto si pensava.
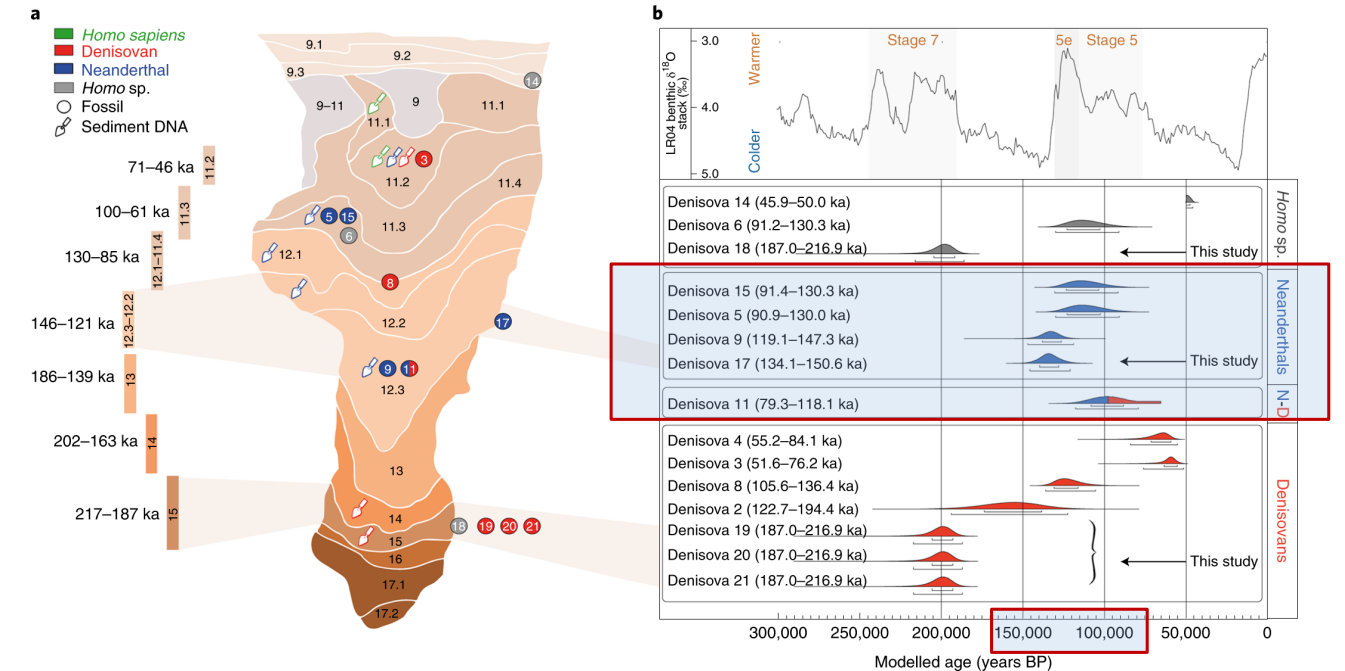
Denisova 11
Denisova 11 è un frammento di femore che presenta sia DNA Neanderthaliano che Denisoviano. E' datato a 90 Ka.
L'individuo ha madre Neanderthal e padre Denisoviano, potrebbe essere stato un evento comune.
Distribuzione
Studi più recenti vedono l'espansione dei denisoviani per anche altre aree dell'Asia. Ci sono tre reperti che metterebbero in luce altri siti dove sono stati rinvenuti reperti attribuibili ai denisoviani. (Nord, Centro Cina, Penghu da Taiwan, Laos.
Il range di distribuzione cronologica è ampio, e pure il range geografico.
La distribuzione del DNA di Denisoviani all'interno delle popolazioni odierne suggerisce che questo gruppo umano fosse ampiamente distribuito in Asia, forse persino fino a circa 25'000 anni fa. Inoltre i bassi livelli di eterozigosi e di consanguineità suggeriscono che avessero popolazioni piccole, ma meno isolate dei Neandertaliani.
Harbin
Il cranio di Harbin era attribuito a H. longi (non tanto condiviso). Da studi su aree del cranio (rocca petrosa dell'osso temporale) in cui si conservano proteine e DNA mitocondriale, si può attribuire in realtà a popolazioni più orientali denisoviane.
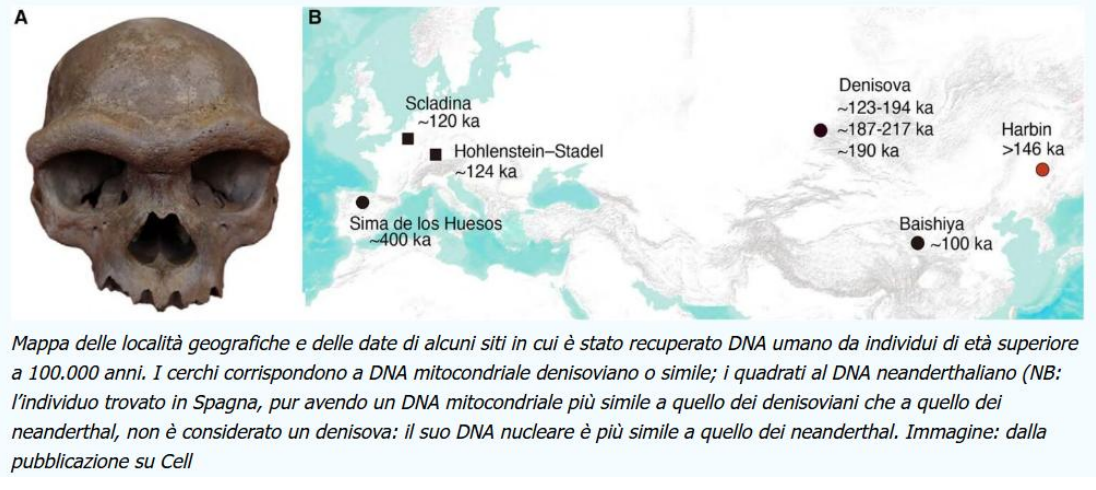
Inoltre, morfologicamente, coincide bene con la mandibola di Xiahe, potrebbe essere questa quindi un reperto denisoviano?
Mandibola Xiahe
Analisi delle paleoproteine ha attribuito la mandibola ai popolazioni vicine ai denisoviani. Altri frammenti sempre di Xiahe sono attribuiti a denisoviani, con datazioni anche relativamente recenti (48-32 Ka)
Penghu Taiwan
Analisi delle proteine lo ha attribuito a un maschio denisoviano, ha una datazione tra i 10-70 Ka o tra i 130-190 Ka. Studio del 2025
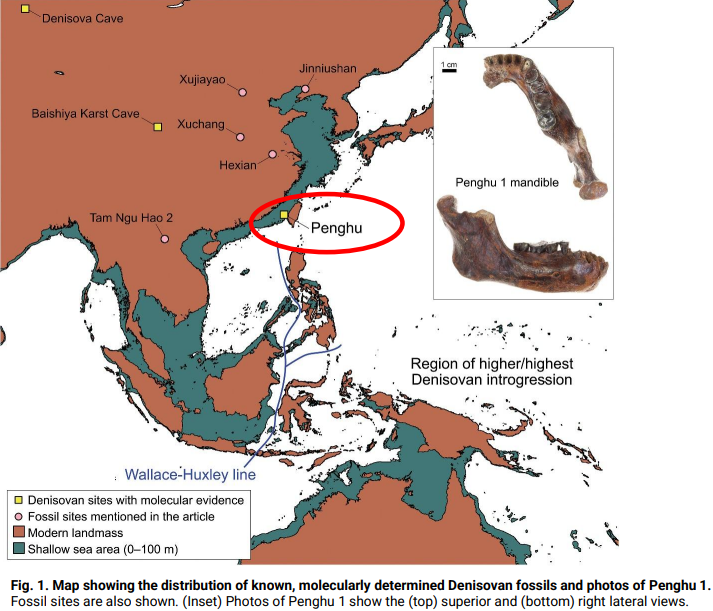
Discendenza denisoviana
E' presente un aplotipo del gene EPAS1, mette in evidenza una ancestralità delle popolazioni denisoviane rispetto alle popolazioni attuali del Tibet. E' un gene che favorisce la resistenza all'ipossia, presente in popolazioni Tibetane e di etnia Han.
Popolazioni in Papua Nuova Guinea mostrano discendenza Denisoviana.
Altre popolazioni con discendenze Denisoviane sono Siberiani e popolazioni dell'America Settentrionale.
Revisione di Bae
E' in corso una profonda revisione di molti resti rinvenuti in Asia.
Ricercatore che propone una revisione dei fossili del Pleistocene medio in Asia orientale.
Attribuisce molti reperti a H. juluensis, una specie molto robusta. Attribuisce diversi reperti (tra cui Harbin) a H. longi (lavoro del 2024, nel 2025 li attribuisce a Denisova).
Un altro è H. luzonensis, nelle Filippine.
H. floresiensis, nell'Isola di Java.
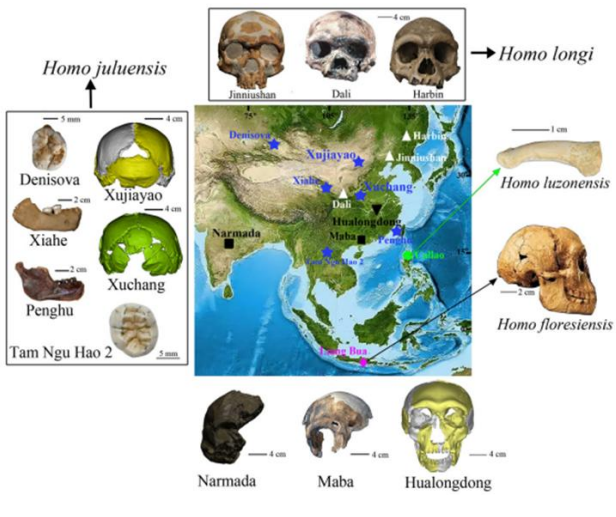
Sommario evoluzione di Neanderthal e Denisova
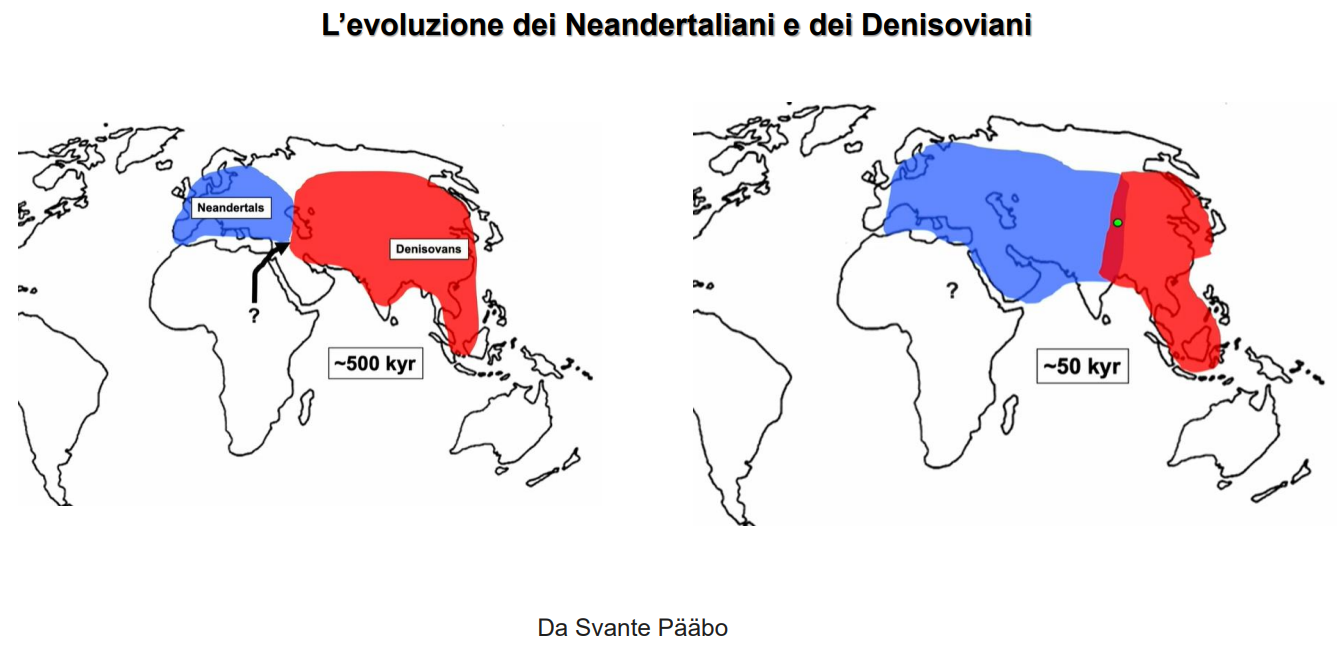
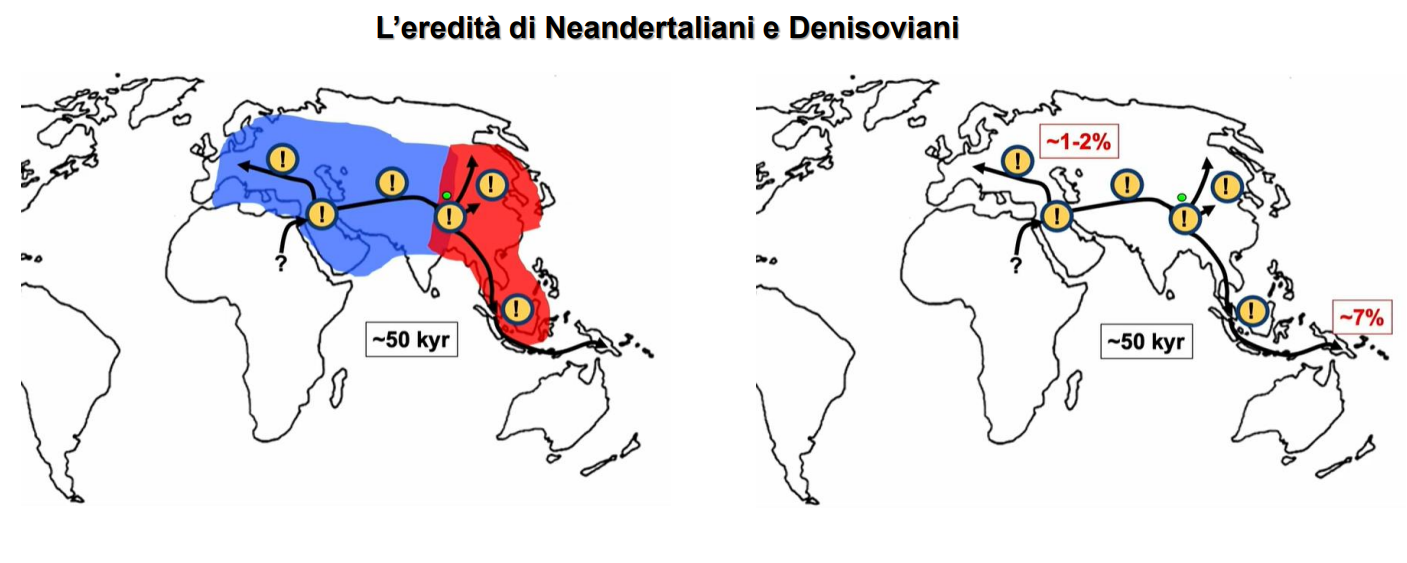
12 Confronto tra caratteristiche biologiche e culturali tra H. neanderthalensis e H. sapiens
Caratteristiche biologiche:
Homo sapiens
Craniale: Capacità cranica di:
Il cranio è globulare.
C'è un rilievo sopraorbitale, non c'è un torus.
Il prognatismo è assente.
Il foto mentoniero è in P2-P1.
Postcraniale:
Culturali:
Le tecnologie litiche di transizione son difficili da attribuire a una specie soltanto siccome sono coeve. Appr.
a Sapiens sono associate le tecnologie litiche xx, e dai 40-39 Ka la tecnologia Aurignaziana,
Il modello demografico è di maggiore successo rispetto a Neanderthal, le popolazioni sono relativamente numerose e interconnesse. C'erano regioni caratterizzate da nuclei, che sostenevano la stabilità nella regione. Tutto grazia a una grande mobilità.
Espansione grazia all'utilizzo dell'ago per cucire, e costruzione di rifugi artificiali.
Ai sapiens sono attestate le realizzazioni di pitture rupestri (Chauvet 32 Ka) e nella cultura maddaleniana anche le pitture del ciclo franco-cantabrico, nelle grotte di Lascaux (17-15 Ka), Altamira (18-14 Ka) , la realizzazioni di Veneri.
Homo neandertalensis
Craniale: La capacità cranica è maggiore di H. sapiens. Il cranio è allungato, non presenta "globularità".
E' presente un torus continuo: la regione sopraorbitale e sopraciliare sono fuse. In sapiens è presente solo un rilievo.
C'è prognatismo medio-facciale (zihomi e mediofaccia spinti in avanti)
E' assente il mento. E' assente la fossa canina.
Il foro mentoniero in Neanderthal è sotto il primo molare (M1), a differenza di sapiens
E' presente la fossa soprainiaca nella parte posteriore.
i denti hanno una superficie ampie e spessore importante, le radici sono molto fuse.
Postcraniale
Il ramo pubico è più lungo e sottile che in sapiens. L'inserzione del muscolo piccolo rotondo avviene su una faccia distinta della scapola rispetto a Sapiens. La sezione laterale del femore è rotonda e non triangolare come in Sapiens
Tronco e bacino più largo.
Le ossa sono molto robuste, cioè ispessite, se comparate a quelle di sapiens.
Piede pronato, piede più piatto.
Caratteristiche culturali:
Le prime sepolture in assoluto sono attestate a Neanderthal. Appr.
La tecnologia litica associata a Neanderthal è il musteriano (Modo 3), perfezionano la tecnica Levallois. (Nei Monti Altai però la tecnologia è ancora indietro, micoquiana)
Le popolazioni erano spesso fatte da piccoli gruppi, relativamente isolati e chiaramente endogamici (Monti Altai).
E' attestato l'utilizzo di ornamenti (Fumane), pigmenti scuri. L'utilizzo del fuoco è attestato. I primi strumenti specializzati in osso sono attestati ai Neanderthal.
13 L'evoluzione di Homo sapiens, le rotte di espansione e il popolamento dell'Eurasia
I mean, vi rimando direttamente qua per l'evoluzione di Homo sapiens.
qua per le rotte di espansione e popolamento dell'Eurasia. (Riguardate la domanda OOA3)
14 La transizione Olocenica - Effetti biologici e culturali
Tredicesima lezione link
C'è da comprendere il contesto culturale antecedente alla transizione Olocenica.
Nella Necropoli di Taforalt (15-13 Ka) in Marocco sono presente numerosi segni di attività culturale. Ci sono elaborati rituali funerari, l'uso dell'ocra con possibili fini simbolici, estensiva manipolazione dei cadaveri e degli scheletri.
Nel Mesolitoco (11-7.5 Ka)
Alla fine dell'ultima era glaciale le comunità paleolitiche si adattano a un nuovo contesto ambientale interglaciale, con aree forestali e un livello del mare alzato.
Si estingue la megafauna pleistocenica. Questo momento mesolitico terminerà con l'arrivo di allevatori e agricoltori dal Vicino Oriente, dove inizia il Neolitico.
Nel Neolitico (<8 Ka) vengono addomesticati diversi cereali, ora integranti per la dieta. Si addomesticano nuove specie, come i grandi mammiferi.
Cambiano anche i temi di certe produzioni culturali e aumentano notevolmente le sepolture, rispetto al Paleolitico, dove erano rare e legate a individui con patologie.
- Aumenta la sedentarietà delle popolazioni
- C'è un effetto di gracilizzazione
- Compaiono le carie (integrazione di molti carboidrati)
- Compaiono stress nel corso dello sviluppo, osteoporosi
Ricerche sulla composizione del microbioma orale (sotto forma di tartaro antico) riflettono questi cambi della dieta (diversi microbi riflettono distinti alimentazioni), dati dall' introduzione dell'agricoltura e la scomparsa delle specie presenti nei cacciatori-raccoglitori.
Si nota inoltre (tramite studi sui siti di Grotta Scaloria e Passo di Corvo) che erano frequenti pratiche funerarie distinte alle semplici sepolture primarie: ciò indicava una frequente manipolazione dei resti umani.
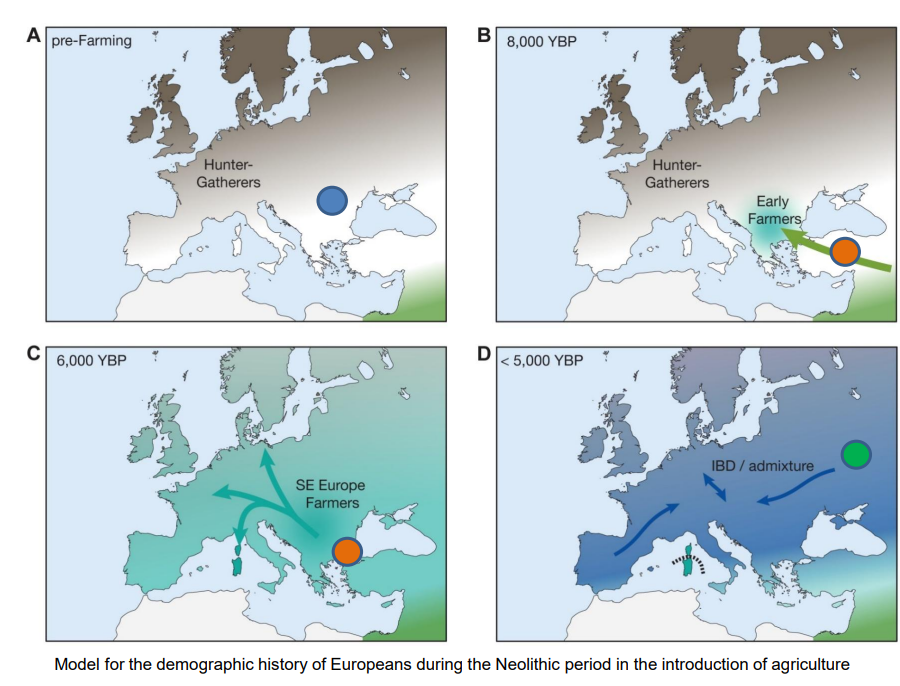
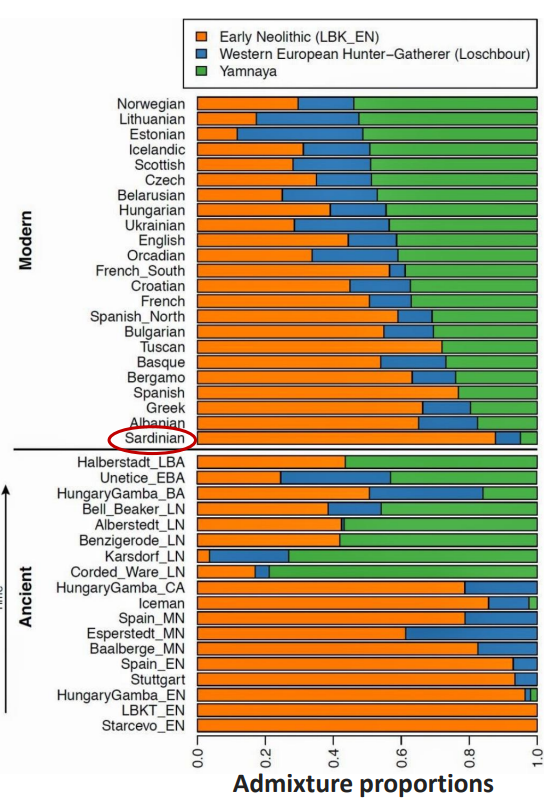
Si denotano tre maggiori contribuenti alla diversità genetica europea. I cacciatori-raccoglitori mesolitici, già presenti da 45 Ka, I primi agricoltori neolitici. arrivati attorno ai 7 Ka, e i pastori delle steppe. arrivati a partire dai 5-4.5 Ka.
Queste ultime culture Yamnaya potrebbero essere stati coloro che hanno introdotto le lingue indoeuropee in Europa. L'arrivo in proporzioni di uomini-donne di 10:1 fa supporre una cultura guerrigliera già sviluppata, grazie anche all'allevamento dei cavalli e la invenzione della ruota e del carro.
Inoltre, potrebbero aver portato il gene che permette la tolleranza del lattosio in età adulta.
15 Elenca i fattori della biodiversità umana
Quattordicesima lezione link
Fattori biologici:
Un fattore fondamentale è la diversità genetica, le informazioni contenute lungo le sequenze di basi nucleotidiche. Questa può interagire con l'ambiente, e venirne modificata, dando nuove informazioni e guidando i meccanismi di sviluppo e acclimatazione (Epigenetica).
Sotto forma di cromosomi, molte di queste informazioni (geni) sono ereditabili nel corso delle generazioni. Tuttavia il genoma viene ricombinato in seguito alla formazione dello zigote (fusione di due cellule aploidi).
Un'eccezione sono il mtDNA (DNA Mitocondriale), ereditato per via materna e la porzione non ricombinante del cromosoma Y, ereditato per via paterna.
Il mtDNA è sprovvisto di introni, ha 37 geni e una zona di controllo. Inoltre ha due zone ipervariabili, con un alto tasso di mutazioni.
D'altra parte il cromosoma Y non ha un propriamente un cromosoma omologo con cui ricombinare, solo il 5% della lunghezza può ricombinare con Y, al fine della spermatogenesi.
L'espressione genetica si riflette sul fenotipo. Ci sono distinte maniere in cui un genotipo può riflettersi:
- dominanza
- recessività
- poliginia (varianti che agiscono in rete con altre varianti)
- pleiotropia (varianti con un effetto su più fenotipi)
Le varianti poi, se funzionali, sono sotto effetto della selezione naturale. Altrimenti passano silenziosamente e rimangono in base alla composizione della popolazione.
Ricorda il linkage (certe varianti possono essere "linkate").
Le varianti possono essere distinte in base alla quantità di genoma interessato
- puntiformi, con diversi effetti se è in una regione codificante e con diverse portate se è una sostituzione o delezione/addizione (frame shift)
- variazioni del numero di copie
- strutturali: Estesi riarrangiamenti (SV o structural variations) dovuti a grandi indels, duplicazioni, inversioni (rottura e riattacco dopo rotazione a 180°), traslocazioni (trasferimento tra due o più cromosomi diversi)
Puntiformi:
I primi cacciatori raccoglitori e i primi agricoltori neolitici non avevano il gene che permetteva la digestione del lattosio in età adulta. Solo 4500 Ka con l'arrivo di allevatori delle steppe questo gene è stato portato.
Inoltre, i primi cacciatori raccoglitori che abitavano la Spagna, Lussemburgo e Ungheria portavano i geni indicatori di pelle scura. In Nord Europa invece, i cacciatori raccoglitori già presentavano geni per la depigmentazione e gli occhi azzurri. L'arrivo dei agricoltori neolitici ha consolidato però la presenza dei geni per la depigmentazione.
Strutturali:
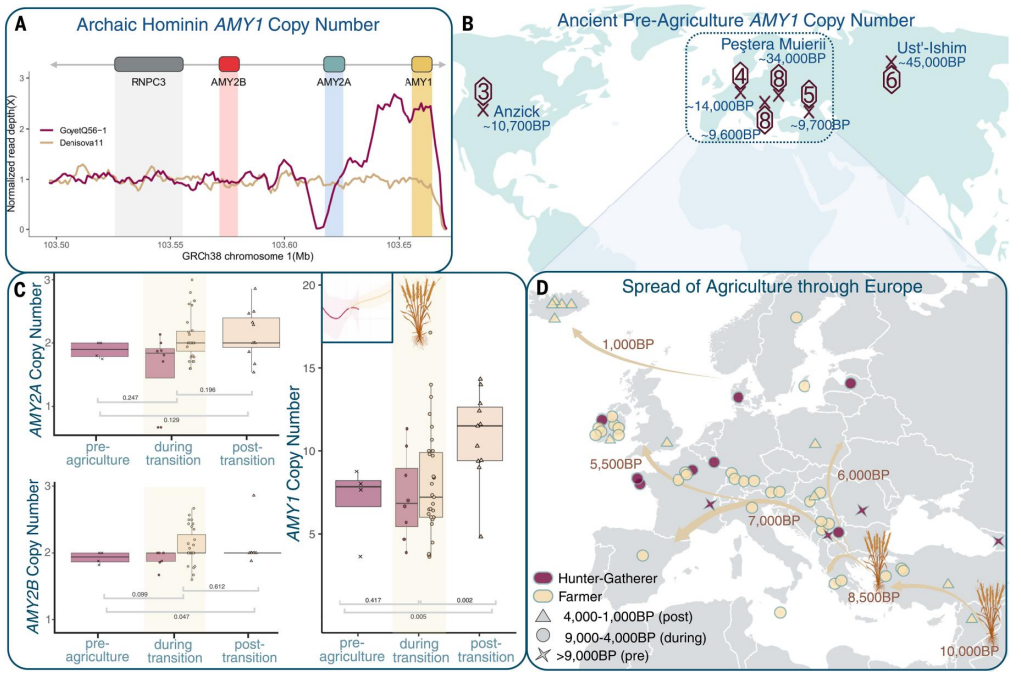
gli antichi cacciatori raccoglitori e persino i Neanderthal avevano molte copie del gene AMY1, che permette la predigestione dell'amido da parte della saliva. Ciò può avere contribuito a cambi importanti della dieta.
Genetica di popolazione
In assenza di fattori esterni, una popolazione rispetta l'Equilibrio di Hardy-Weinberg. Implica un'assenza di cambiamenti e quindi di evoluzione.
Le deviazioni da questo modello sono le forze evolutive che hanno causato cambiamenti nel tempo.
Molte di queste non sono adattative, causano cambiamenti e variabilità ma non necessariamente influenzano la capacità di un individuo di sopravvivere a riprodursi. Sono Inbreeding, Migrazioni, Deriva genetica e Isolamento. La Selezione naturale invece è l'unica forza evolutiva adattativa.
Inbreeding:
Può essere casuale (quindi c'è un livello atteso in base alla dimensione della popolazione) o non casuale (se il partner viene selezionato in base al livello di parentela). All'aumentare dell'inbreeding si assiste a un calo dell'eterozigosità, con diverse conseguenze: perdendo variabilità si perde resistenza a malattie, fertilità...
Isolamento e Deriva genetica:
Se avviene un evento che isola una parte della popolazione, si possono fissare in una nuova popolazione varianti alleliche non rappresentative della popolazione iniziale. Gli effetti della deriva sono maggiori più è piccola la popolazione.
La deriva caratterizza popolazioni distinte, diminuisce la diversità interna a una subpopolazione e aumenta la diversità tra le popolazioni.
Migrazione:
Le migrazioni hanno effetto opposto alla deriva, normalizzando le differenze tra metapopolazioni. Tuttavia queste avvengono in base alla distanza delle popolazioni, le somiglianze quindi si notano su un gradiente
La migrazione può essere sex-biased*, su piccola scala spesso è la donna a migrare verso la unità sociale del marito (patrilocalità). E' confermato nei Monti Altai: il gruppo era relativamente isolato, ma collegato con altri per migrazione femminile.
Ciò causa una omogeneizzazione del mtDNA su piccola scala. Su grande scala invece le migrazioni sono spesso sex-biased per i maschi.
Selezione naturale:
La selezione avviene sulle informazioni genetiche (sul patrimonio genetico) ed epigenetiche (sui meccanismi di controllo ed espressione del patrimonio genetico, senza mutarlo).
Selezione negativa: Agisce sulle aree codificanti, ed elimina le varianti deleterie.
Selezione bilanciata: Il vantaggio è dato dall'eterozigosi, da più alleli del locus.
Selezione positiva: possono esserci grandi eventi di selezione (Hard selective sweep), che riducono la variabilità del genoma selezionato. Per effetto del linkage possono essere selezionati anche tratti sullo stesso cromosoma, non adattativi.
Esempi di questo sono stati la selezione per la pelle chiara alle latitudini più a nord, e la tolleranza del lattosio.
Introgressione adattativa: è l'introduzione di informazioni genetiche adattative da un'altra specie interfeconda. Un esempio è l'aplotipo del gene EPAS1 che permette un adattamento alle altitudini, nelle popolazioni Est-Asiatiche, portato dai denisoviani.
Inoltre, ci sono le regole Ecogeografiche di Allen-Bergmann
Gli organismi endotermici in aree fredde tendono ad aumentare la massa corporea e le dimensioni, così da disperdere meno calore (ratio superficie/volume minore), inoltre le appendici tendono ad essere più contenute.
