Homo habilis e rudolfensis
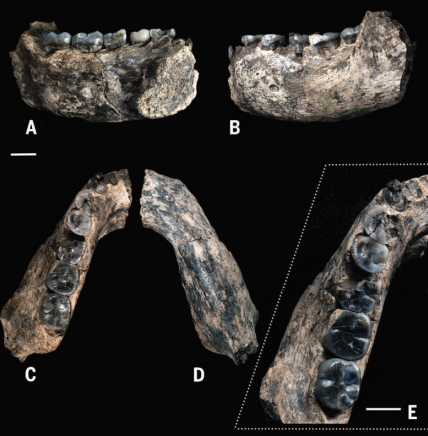
Dalle slide, pare che si sia una mandibola ancora più antica attribuita a un possibile antenato di H. habilis e rudolfensis, di 2,8 Ma. L'arcata è molto larga, a dimostrare una grande varietà ortognatica dei Early Homo.
habilis
Range 2,3 Ma - 1,5/1,8 Ma
La specie è stata descritta sulla base di reperti datati a 1,8Ma. Nello stesso sito sono stati ritrovati anche fossili attribuibili a P. boisei, ma le morfologie del cranio era ben distinguibile.
E' la prima inequivocabile evidenza della coesistenza di due specie di Ominini.
I reperti più antichi sono stati rinvenuti ad Hadar in Etiopia ed attribuiti a 2,3Ma. Altri resti sono stati rinvenuti anche in vari siti in Kenya, Tanzania e persino in SudAfrica.
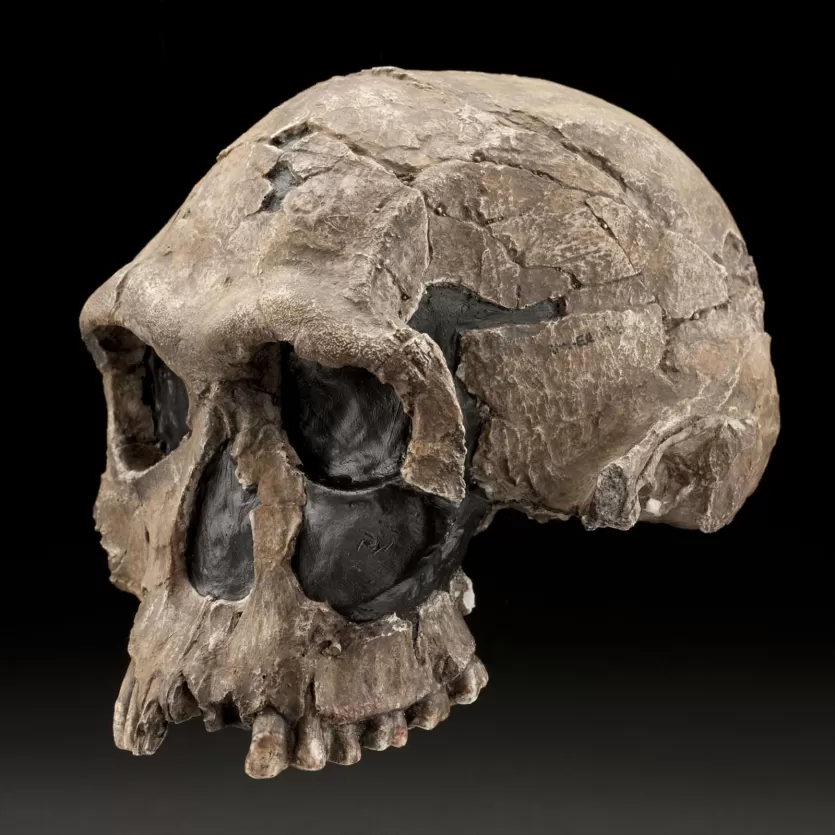
Importante è un cranio rinvenuto in Kenya, datato a 1,9Ma a Koobi Fora nei pressi del Lago Turkana.

La struttura è complessivamente simile ai A. africanus, ma si distingue per lo sviluppo dell'arcata sopraorbitaria, ossa nasali prominenti e dimensioni dentarie più piccole.
In generale H. habilis la capacità cranica varia dai 500 ai quasi 800cc, principale differenza rispetto ai Australopitechi.
Sebbene ci siano pochi resti post-craniali attribuiti, suggeriscono una struttura e proporzione degli arti tale per cui è mantenuta una capacità di arrampicamento.
Assieme a H. rudolfensis, pare che la struttura postcraniale fosse sostanzialmente simile a quella degli australopitechi, quindi l'andatura bipede potrebbe essere stata facoltativa. Non sono ancora presenti le strutture dello scheletro che accomuneranno tutte le specie del genere Homo successive, sono più simili a "Australopitechi con cervello di dimensioni più grande". Questa ipotesi di Ominini di "transizione" lascia aperta l'ipotesi che non siano nemmeno ascrivibili al genere Homo.
Homo rudolfensis
Range: 2,5 Ma - 1,6 Ma
E' un'altra specie alla base del genere Homo, di cui però si conoscono pochi resti craniali e dentari, sono assenti resti post-craniali.
La capacità cranica è maggiore (760-820cc), e manca il toro sopraorbitario. La faccia ha un prognatismo ridotto. I denti sono di dimensioni maggiori rispetto a quelli di H. habilis e più simili a quelli dei Paranthropus.
La diversità delle morfologie fa supporre un adattamento a diete distinte, siccome comunque erano specie coesistenti negli stessi areali.
E' molto discussa la attribuzione di questi reperti, se costituisce o meno una specie a se stante (o è parte del dimorfismo di H. habilis), o a che genere appartiene (inizialmente è stato descritto come appartenente a Pithecanthropus). E' inserito nel genere Homo per la elevata capacità cranica.

Da Wikipedia:
Beyond KNM-ER 1470, there is disagreement on which specimens actually belong in H. rudolfensis as it is difficult to assign with accuracy remains that do not preserve the face and jaw. No bodily elements have been definitively associated with a skull and thus to the species.
Most proposed H. rudolfensis fossils come from Koobi Fora and date to 1.9–1.85 mya. Remains from the Shungura Formation, Ethiopia, and Uraha, Malawi, are dated as far back as 2.5–2.4 mya, which would make it the earliest identified species of Homo. The latest potential specimen is KNM-ER 819 dating to 1.65–1.55 mya.
Evidenze culturali
Ciò che ha determinato la attribuzione al genere Homo di queste due specie è il rinvenimento di utensili di pietra in depositi coevi.
Però studi più recenti che dimostrano la presenza di utensili in depositi più antichi pare escludere l'idea che la costruzione di utensili fosse esclusiva del genere Homo.
Tuttavia le evidenze più definite e numerose sono coetanee a queste specie del genere Homo.
La creazione di semplici utensili in pietra implica numerose specializzazioni e novità. Grazia all'andatura bipede gli arti superiori sono svincolati dalla necessità di sostegno del corpo.
Inoltre la realizzazione di questi utensili richiede lo sviluppo di specifiche aree cerebrali così da poter dare la coordinazione e finezza necessaria
In questo periodo sono pure attestate le prime evidenze di destrismo
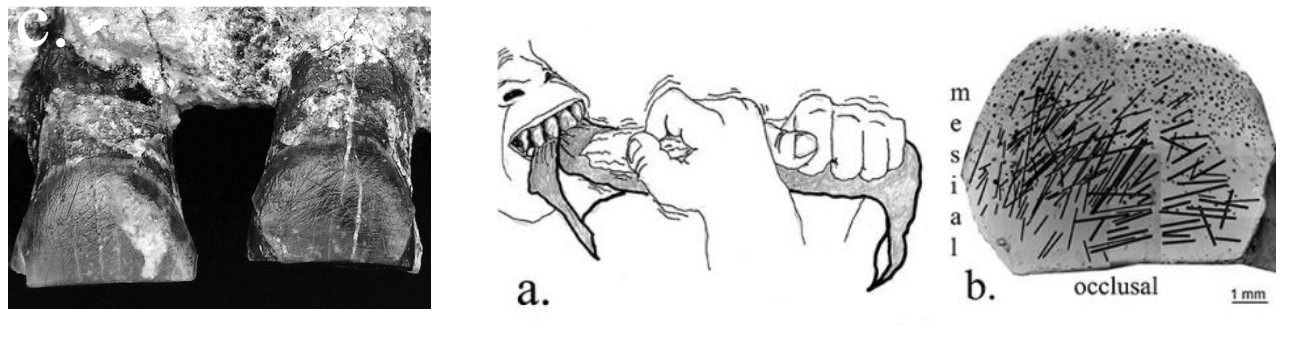
e i primi interventi intenzionali su resti umani
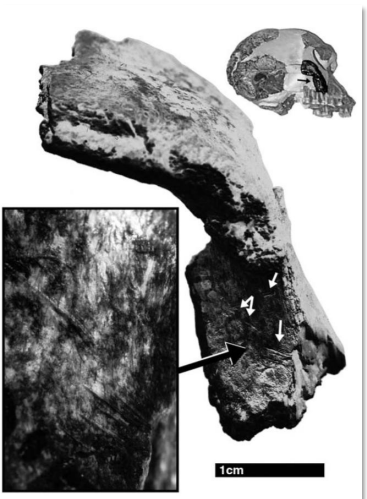
E' ancora discusso inoltre il ruolo della carne nella dieta di questi ominidi, sono state ritrovate ossa di grandi mammiferi con evidenti segni di scarnificazione, ma non è chiaro se fossero stati cacciati da questi ominidi o se fossero carcasse a cui hanno avuto accesso.
Tuttavia l'accesso alla carne deve aver dato l'apporto calorico a lungo termine necessario allo sviluppo del cervello
Homo erectus s.l
Erectus s.l
Caratterizza il Pleistocene inferiore.
E' probabile che sia originato da alcune popolazioni arcaiche africane di H. habilis entro le quali sono avvenuti eventi di speciazioni, che hanno dato origine alle forme che oggi identifichiamo come H. erectus e H. ergaster
Se di H. habilis e H. rudolfensis non è chiara l'appartenenza al genere Homo, per H. erectus non ci sono dubbi.

E' una specie che si è diffusa ampiamente, dal SudAfrica al Sudest asiatico, in un lasso temporale molto largo, dai 2Ma per le forme più antiche fino ai 117'000 e 108'000 anni fa.
I primi reperti sono stati trovati sull'Isola di Java, ed il reperto più antico è stato trovato in SudAfrica.
Ciò ha portato a una grande varietà morfologica, tale per cui molti autori considerano H. ergaster come una specie separata (forma africana), mentre H. erectus sensu strictu come la forma diffusa in Asia.
Entrambe le forme (da qui H. erectus s.l.) mostrano una chiara discontinuità con le forme precedenti e molte morfologie derivate.
La morfologia del cranio è allungato in senso antero-posteriore, con una marcata protuberanza dell'osso occipitale, detto Toro occipitale, le arcate formano un Toro sopraccigliario. Presenta una protuberanza lungo la sutura saggittale (non una cresta come in Paranthropus), chiamata anche carena occipitale.
La fronte è molto sfuggente.
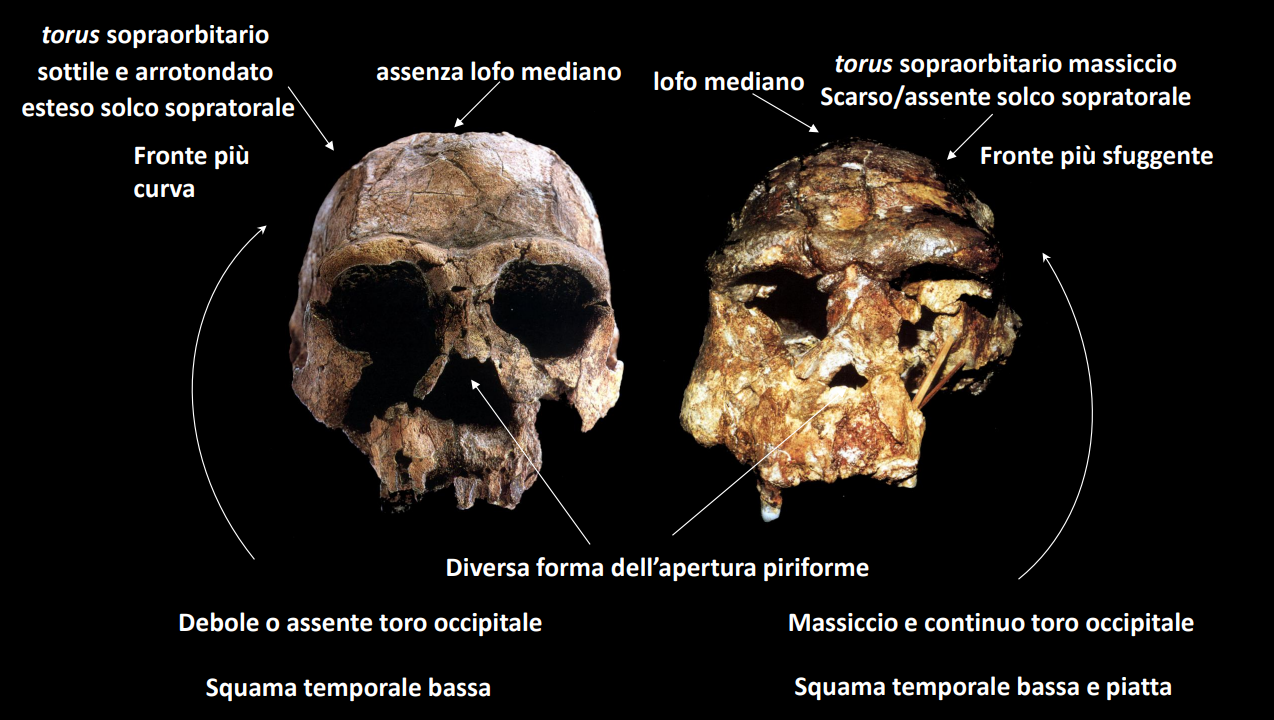
Sx H. ergaster, Dx H. erectus di Giava
La capacità cranica media della specie s.l. è superiore rispetto a H. habilis e H. rudolfensis, con valori medi attorno ai 800cc per le forme africane e 980cc per le forme asiatiche.
Inoltre una dieta chiaramente distinta, con un maggior apporto di cibi ricchi dal punto di vista energetico come le proteine animali, hanno determinato l'avvio di una diminuzione della superficie masticatoria dei denti.

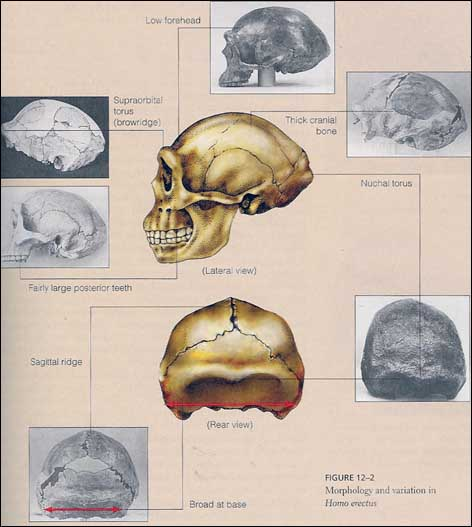
I resti postcraniali di H. erectus mostrano inoltre che questa specie deve aver perso la capacità di arrampicamento sugli alberi, che ancora caratterizzava H. habilis.(L'articolazione della spalla non è più rivolta cranialmente ma lateralmente)
La struttura del bacino e dell'arto inferiore mostrano adattamenti a una locomozione bipede esclusiva. Ad esempio il femore è più allungato e proporzionalmente gli arti inferiori sono più lunghi rispetto ai superiori, l'arco plantare è ben sviluppato e l'alluce è completamente parallelo alle altre dita del piede.
E' una morfologia del piede chiaramente moderna, anche se non è chiaro se potesse correre su lunghe distanze come l'essere umano moderno.
Homo ergaster
A H. ergaster vengono attribuite i primi resti del genere Homo trovato al di fuori dell'africa Out of Africa
Da questo, evoluto circa 2 Ma. Avvengono le prime dispersioni out of Africa.
Già sviluppa la tecnologia litica
Ragazzo del Turkwana
Molto informativo è stato il ritrovamento dei resti di un individuo, la cui completezza è attorno all'80%. E' stato rinvenuto in Kenya ed è risalente attorno ai 1,5Ma.
E' attribuito H. erectus s.l.. Tuttavia essendo un subadulto non fornisce un'immagine degli individui completamente cresciuti.
Ha molte caratteristiche affini a H. sapiens e rappresenta un chiaro momento di stacco con le forme precedenti.
Probabilmente sudava come l'essere umano moderno, quindi non era dotato di una folta peluria. E studi della pelvi mostrano come non fosse sufficientemente larga da lasciar passare un grande cranio durante il parto, perciò i neonati dovevano nascere prima che l'encefalo fosse completamente formato, come nell'essere umano moderno.

Impronte di Ileret
Sono impronte attribuite a H. ergaster, datate tra i 1,53-1,51 Ma. Mostrano che gli ominini del genere Homo avevano già un piede morfologicamente moderno, con pollice addotto.

Manifestazioni culturali
Con H. erectus s.l. si manifestano anche tradizioni culturali innovative, come la cosiddetta cultura Aucheuleana, caratterizzata da bifacciali o amigdale, che però non arriva in Asia.
Questa specie sicuramente praticava una caccia attiva e non solo sciacallaggio, grazie ai nuovi utensili e la crescente capacità cranica.
Anche in Asia ci sono state distinti eventi pleistocenici di diffusione Out of Africa. I più antichi di questi eventi potrebbero aver dato poi origine a forme morfologicamente arcaiche come H. floresiensis.
E' molto documentata la presenza in Estremo Oriente di H. erectus s.s. Nel Pleistocene medio si rinvengono fossili in siti cinesi e indonesiani.
Morfologicamente è molto caratterizzante la presenza di una volta bassa che con una prospettiva laterale comporta un profilo posteriore "a cuneo".
Al contesto asiatico è associata una tecnologia litica modo 1.
Pleistocene medio
Il Pleistocene medio è un importante periodo di transizione. Verso la fine vediamo la comparsa della nostra specie, assieme a quella dei Neanderthal e dei Denisoviani. Queste tre specie sono originate da un antenato comune, H. heidelbergensis. Nel contempo coesistono altre specie, come H. erectus nel Sudest asiatico, H. naledi in Africa e H. floresiensis nell'Isola di Flores (nell'arcipelago della Sonda). Anche H. antecessor in spagna.
In questo periodo le varie specie mostrano la transizione verso forme più derivate, pur mantenendo morfologie riferibili a H. erectus s.l.. C'è una tendenza verso una maggiore encefalizzazione e lo sviluppo di nuove tecnologie, come il fuoco e la tecnica Levallois.
La complessità tassonomica si può relazionare all'instabilità climatica che si instaura nel passaggio dal Pleistocene inferiore al Pleistocene medio.
La ciclicità dei climi passa dai 40ka ai 100ka, quindi le differenze si fanno più marcate tra momenti caldo-umidi (temperati al nord) e secco-freddi (glaciali al nord). Questa forte eterogeneità degli ambienti e dei climi ha fortemente marcato l'evoluzione degli ecosistemi.
Homo heidelbergensis
heidelbergensis
Evoluto circa 600 ka fa da Homo ergaster, in Africa
Ha caratteristiche tali per cui viene richiesta la definizione di una specie a se stante, capacità cranica (1000cc) e statura molto piccola.
Darà origine a diverse specie Da origine a Homo sapiens e dall'altra parte in Europa e Asia altre forme di heidelbergensis(con bifacciale), da cui origineranno Homo neanderthalensis (200ka, 1400cc) e l'uomo Denisoviano
Nasce in Africa.
Grande variabilità di forme e morfologie, originata dalla variabilità già presente in H. ergaster, però tutte accomunate da una crescita dell'encefalo dai 800cc fino anche ai 1200cc (già notabile in H. ergaster).
Attorno ai 600'000 anni fa un gruppo di H. heidelbergensis esce dall'africa.
I primi manufatti litici di tipo x sono stati trovati già 700'000 anni fa, mentre i primi resti umani a partire dai 500'000 anni fa.
H. heidelbergensis deve essere stato un taxon molto variabile morfologicamente e di ampia distribuzione geografica e deve essere vissuto per tutto il Pleistocene medio, tra i 600Ka e i 200Ka.
Tale processo deve essere legato alla fuoriuscita dall'Africa di popolazioni di cacciatori raccoglitori caratterizzate da Industrie litiche di Modo 2, che devono aver soppiantato popolazioni ancora di Modo 1.
.
Homo heidelbergensis esiste come specie ancestrale, variabile al suo interno con presenza di popolazioni semi-isolate (paleodemi o sottospecie), antenata di H.n., H.s. e probabilmente Denisoviani che si differenziano dal gruppo ancestrale e tra loro, evolvendosi in specie diverse per ragioni di ordine geografico-ambientale-demografico
Ipotesi alternative - Homo antecessor
Per alcuni studiosi la variabilità di H. heidelbergensis è eccessiva per poter essere considerato un taxon unico. Le distinte popolazioni potevano essere già sufficientemente derivate così da distinguerle nei distinti taxa (non sono quindi forme di H. heidelbergensis, ma forme antiche delle specie da questo derivate).
In Europa si riscontrano i cosiddetti Early neanderthal (reperti con caratteri mosaici ascrivibili a forme arcaiche, ma anche novità che saranno caratterizzanti dei Neanderthal). In Asia ci rinvengono forme già associabili ai Denisova, mentre in Africa appaiono cambiamenti "moderni", come la modificazione della faccia e della dentatura, e dai 200'000 anni l'acquisizione della globularità del cranio.
La specie ancestrale alternativa potrebbe essere quindi stata il cosiddetto H. antecessor, rinvenuto soltanto in Spagna a Sierra de Atapuerca*, datato a 800Ka, con una formazione "moderna" della regione mascellare.
Però a supportare l'ipotesi che H. heidelbergensis sia antenato comune delle specie venute dopo c'è il ritrovamento di tre mandibole rinvenute nella **grotta di Arago (Francia)*, che mostrano affinità come altri resti di H. heidelbergensis rinvenuti in Algeria, mentre non sono filogeneticamente riconducibili a H. antecessor
Record fossile in Europa
Diversi ritrovamenti, come le mandibole di Arago (Francia) e Mauer non mostrano tratti derivati neanderthaliani.
Ciò non si può dire per i fossili rinvenuti a Sima de los Huesos, con cui però comunque condividono un modello strutturale.
I resti di quest'ultimo sito hanno un mosaico di caratteristiche già riferibili ai Neanderthal, seppur non al punto di essere riferibili come tali.
Ciò fa ipotizzare un'origine anagenetica secondo un modello accrezionale.
Vedi "comparsa della specie" per approfondire.
Resta di fatto che assieme a questi reperti che mostrano un progressivo accumulo di tratti "Neanderthal like", si ritrovano resti che non rispecchiano queste caratteristiche, mantenendo attivo il dibattito sulla questione dell'ancestralità di H. heidelbergensis per i neanderthaliani (I reperti di Sima de los huesos sono già da considerare Neanderthal quanto tali?)
Record fossile in Asia
Mentre era comunque diffuso H. erectus s.s in tutta l'Asia, a introdurre una industria litica modo 2 è stato l'arrivo di H. heidelbergensis a partire da 400Ka.
Questa nuova umanità va a sostituire le popolazioni di H. erectus nell'area continentale. Le specie relitte sopravvivono solo nel sudest asiatico.
Anche qui come in Europa si incontra un contesto morfologico molto variabile, con alcune forme associate a specie tutt'ora dibattute, come H. longi. Questa varietà potrebbe essere alla radice dei denisoviani.
Record fossile in Africa
L'Africa è il continente dove è originato H. sapiens prima di diffondersi nel resto del mondo, ma è anche il continente dove il nostro progenitore, H. heidelbergensis si è originato.
C'è una grossa lacuna nella documentazione fossile nell'intervallo compreso tra i 900 e 600 Ka. Le morfologie riconducibili a H. ergaster scompaiono intorno a 1Ma.
Dopo i 600Ka la documentazione fossile torna ad essere abbondante, soprattutto in Africa SudOrientale, con rappresentazioni di caratteristiche derivate, nonostante siano mantenuti tratti arcaici.
- Profilo frontale arcuato
- Volumi cerebrali medi di 1200 mL
I reperti vengono spesso, ma non unaminimamente, attribuiti a H. heidelbergensis.
A partire dai 300Ka si trovano morfologie ancora più derivate, con volumi cerebrali maggiori, taglia minore di denti e faccia, riduzione del toro sopraorbitario.
Alcuni autori attribuiscono questi reperti a una nuova specie, denominata H. helmei.
A queste forme si associa una cultura litica più complessa, Modo 3 con la lavorazione su scheggia
Manifestazioni culturali
Diversi reperti culturali attribuiti a H. heidelbergensis
- Abbiamo indizi di utilizzo di Ocra rossa (250'000).
- E' stata trovata una tibia di elefante a Bilzinsgleben, dove vennero ritrovati diversi segni, difficilmente riconducibile a danni chimici/ambientali. E' plausibile un'origine umana, possibili catmarks, causati dal ripulire le ossa, scotennare l'animale
- Tra 400'-200'000 anni fa è stato ritrovato un bifacciale con anche un valore estetico (conchiglia)
- Legni induriti dal fuoco, utilizzati per la caccia throwing sticks. Importante ritrovamento perchè il legno è di difficile conservazione.
- Conchiglia rinvenuta a Java 430'000. Evidenza dell'utilizzo dei bivalvi a scopo alimentare. Con un motivo particolare come segno di una possibile manifestazione cognitiva.
- Inizialmente attribuito a H. erectus, ma potrebbe essere attribuito a H. heidelbergensis
- Le prime veneri. dai 300'000 anni fa, in Palestina e in Marocco.
Homo floresiensis
floresiensis
Ritrovato nell'isola di Flores a Liang Bua, attribuito al genere di Homo. Con però molte anomalie (cranio molto piccolo, statura molto ridotta).
Si ipotizza sia un caso di nanismo insulare.
Hanno datazioni molto recenti, 90/50'000.
In anni più recenti, è stato trovato sempre a Flores un frammento di mandibola, sempre attribuibile a H. floresiensis, datato però a 1mln-700'000 anni fa. Non è chiarissimo se è attribuibile a H. erectus
Origine di nuove specie
Dal Pleistocene medio provengono le più antiche sequenza paleogenetiche estratte da fossili del genere Homo. Con queste è possibile ricostruire le relazioni filogenetiche tra le varie specie e comprendere quando sono apparse e differenziatesi le nuove linee evolutive.
La separazione tra la linea di Homo neanderthalensis e Homo sapiens è avvenuta circa 500Ka. La separazione con la linea dei Denisoviani pare essere antecedente (600Ka) se si considera il DNA antico, ma il DNA mitocondriale fornisce l'ipotesi che sia stata contemporanea alla separazione dei neanderthal.
C'è da considerare che questi sono i momenti di separazione tra due linee evolutive, ma non corrispondono a eventi di speciazione, che possono essere avvenuti accumulando caratteristiche nel corso del tempo, differenziandosi progressivamente dall'antenato comune e nelle rispettive aree geografiche
Resta molto dibattuto in quale punto le nuove specie si possano considerare come tali e le modalità evolutive che hanno portato alle speciazioni, così come la classificazione delle forme intermedie.
Tra i 150-50'000 anni compaiono nuove forme da H. heidelbergensis, tra cui Homo neanderthalensis, in Europa, Caucaso, vicino oriente, Uzbekistan e Siberia. Non è riconosciuto da tutti il passaggio da H. heidelbergensis a H. neanderthalensis, siccome si ritiene che la capacità cranica fosse già sufficiente da ritenerlo tale come neanderthalensis prima dell'arrivo in Europa.
Poi i Denisoviani, rispettivamente europeo e asiatico. Si estingueranno intorno ai 50'000 anni fa. In questa fase tra Pleistocene medio e superiore, vediamo quindi un grande mescolamento di specie e popolazioni.
Pleistocene superiore
E' un periodo compreso tra i 126'000 e 11'700 anni fa. Anche qui ci sono state importanti oscillazioni climatiche, che hanno visto l'alternanza di periodi glaciali e periodi più umidi. Importante è l'Ultimo massimo glaciale, avvenuto tra i 26'000 e 17'000 anni fa.
All'inizio del Pleistocene superiore, la nostra specie aveva colonizzato già tutte le aree accessibili del continente africano. Grazia all'abbassamento del livello del mare nei periodi freddi, l'essere umano aveva raggiunto la penisola arabica.
Nel Pleistocene superiore L'Eurasia superiore era già occupata da numerosi gruppi umani con cui H. sapiens ha potuto stringere dei rapporti.
Homo sapiens
Evoluto...
La tecnologia prosegue pressochè ininterrottamente. Aiuta a seguire il percorso evolutivo delle popolazione. Viene utilizzato il bifacciale nelle prima fasi dell'homo sapiens. Viene la tecnologia litica modo 3, detta Tecnica Levallois, circa 300.000 anni fa, in Marocco (Jebel Hirhoud) e in Africa Orientale
Jebel Hirhoud
Sempre nel sito di Jebel Hirhoud sono stati trovati resti di ominidi che già mostrano caratteristiche tipiche di Homo sapiens, perchè mostrano nello scheletro facciale (meno in quello neurale) caratteristiche che rientrano tra quelle di Homo sapiens.
I reperti di Homo sapiens hanno la forma globulare del cranio, ed è un carattere diagnostico.
I reperti di Jebel hirhoud sono pre-sapiens, con caratteristiche molto importanti, ma assenti della globularità del cranio.
Omo Etiopia
In Etiopia manca lo scheletro facciale, ma abbiamo il neurocranio, 233'000 anni fa. Ma la globularità è già molto marcata. Con circa 1350cc di volume
Rispetto alle forme arcaiche quindi, la fronte è più verticale, l'occipitale è più globulare, faccia ridotta.

Herto Bouri
Uno dei rappresentanti più noti di Homo sapiens in Africa è il reperto ritrovato in Etiopia a Herto Bouri, che mostrano le caratteristiche tipiche di Homo sapiens. Con una datazione di 150/160'000 anni fa. Ora abbiamo datazioni più antiche (Jebel hirhoud) in seguito a diverse revisioni e scoperte.
Rappresenta l'antichità e l'origine di Homo sapiens in Africa.
Oggi l'emersione di homo sapiens risulta più complessa di quanto si riteneva nel 2003 (data di scoperta di questo reperto). Ma si ritiene appartenenesse a quelle popolazioni che poi si son spostate in Eurasia (grazie a studi delle tracce morfologiche e tecnologia litica).
Questo reperto ha valori di capacità cranica importanti (1450cc), e rappresenta:
forme arcaiche cronologicamente e morfologicamente (Globularità del cranio) che precedono quelle moderne delle ultime fasi del Pleistocene. Rappresenterebbero gli antenati della forma anatomicamente moderna (Anatomical Modern Humans – AMH)
Si ritiene ci fosse un'attività xx, con asportazione del cuoio capelluto, disarticolazione della mandibola. Probabilmente possono essere premesse di tracce di cannibalismo. Reperto di un cranio infantile.
Ipotesi evolutive
Non è chiarissima l'origine di Homo sapiens all'interno dell'Africa: non è chiaro dove sia avvenuto esattamente e che specie gli abbia dato origine (H. heidelbergensis? H. rhodesiensi? H. antecessor).
Esistono diverse ipotesi, che sostengono una possibile origine in Africa Orientale (sulla base di reperti ritrovati in questo contesto, o In SudAfrica, data la diversità di alcune attuali popolazioni di cacciatori-raccoglitori.
Un lavoro recente però ipotizza un'origine panafricana di Homo sapiens, retrocedendo la sua origine tra i 350 e 300Ka, grazia a nuove datazioni del sito di Jebel Hiroud. Mix di caratteri in diversi in diversi siti. Evento anagenetico, graduale, con il contributo di molte popolazioni.
Oppure un evento di speciazione (cladogenetico e allopatrico) a carico di una piccola popolazione di Homo sapiens