Un sistema sensoriale è formate da sensori, da fibre afferenti (da periferia SNC) e aree del SNC che elaborano le informazioni.
Possiamo distinguere tra
- Sistema somato-sensoriale, che riguarda il corpo, superficie e organi interni. Tatto, pressione, temperatura, dolore. I ricettori sono diffusi.
- Sensi speciali: Olfatto, gusto, vista, udito, equilibrio. Con organi specializzati e strutture sensoriali.
Una sensazione è la consapevolezza di uno stimolo, la percezione è il processo attivo per cui si distingue una sensazione tra le altri.
Abbiamo molti tipi di recettori:
- fotorecettori (visione)
- chemiorecettori (olfatto, gusto e dolore)
- recettori termici
- meccanocettori (tatto, udito, equilibrio, propriocezioni)
I recettori normalmente sono collocati su terminazioni libere, sui dendriti. Oppure possono essere cellule modificate, ciliate (udito e equilibrio), che sono in grado di trasdurre un segnale.
Vie afferenti
A questi sistemi fanno eccezione olfatto e vista.
Normalmente hanno un neurone di 1° ordine, dalla periferia al midollo spinale / tronco encefalico; dove fa sinapsi con un neurone di 2° ordine. Il neurone di 2° ordine ascende fino al talamo. Questa stazione talamica è obbligatoria a tutti i sensi, tranne l'olfatto. Dal talamo il neurone proietta alla corteccia cerebrale, tramite un neurone di 3°ordine.
Questo è ciò che ci fa percepire uno stimolo grezzo, ma non è l'unica via. Alla corteccia
primaria arriva lo stimolo grezzo.
Un campo recettivo è l'area in cui uno stimolo può essere percepito. E' dato da neurone, dendriti e tutti i recettori. E' definita unità sensoriale. Ciascuna sensazione ascende al SNC tramite vie afferenti specifiche. Gli stimoli nervosi di origine tattile hanno vie riservate e non si mischieranno mai a quelle uditive.
L'organizzazione topografica è importante, recettori adiacenti proiettano in aree adiacenti a livello della corteccia cerebrale.
I sistemi sensoriali codificano per quattro aspetti di uno stimolo
- Tipo (modalità)
- Intensità
- Provenienza
- Durata
Modalità
Uno stimolo che attiva il recettore con il più basso livello di energia è chiamato stimolo adeguato.
Sono specifiche le vie afferenti e la corteccia a cui proiettano quelle vie: ad esempio strofinarsi l'occhio produce comunque uno stimolo visivo. Il cervello interpreta la via che è stata utilizzata, non è tanto importante il recettore di per sè.
Codifica della localizzazione dello stimolo
Nel caso del sistema somato-sensoriale ci sono aree della corteccia che codificano per ogni area del nostro corpo. Riesco a localizzare uno stimolo somato-sensoriale siccome i neuroni di 3° ordine proiettano su regioni adiacenti.
Nel caso della vista c'è un neurone di 3° ordine che riproduce il campo visivo a livello della corteccia. E' prevista una localizzazione spaziale.
Olfatto e gusto prevedono una codifica di tipo chimico, quindi non è prevista una spazialità.
Nel caso dell'udito i recettori sono localizzati.
Localizzazione dello stimolo: acuità tattile
L'acuità tattile è la capacità di discriminare due punti separati tra loro e sottoposti a stimolo pressorio. Dipende dall'ampiezza dei campi recettivi e dalla quantità.
C'è una soglia di discriminazione tra le parti del corpo. Varia nelle diverse aree della superficie del corpo.
Nelle labbra abbiamo la massima acuità (1mm).
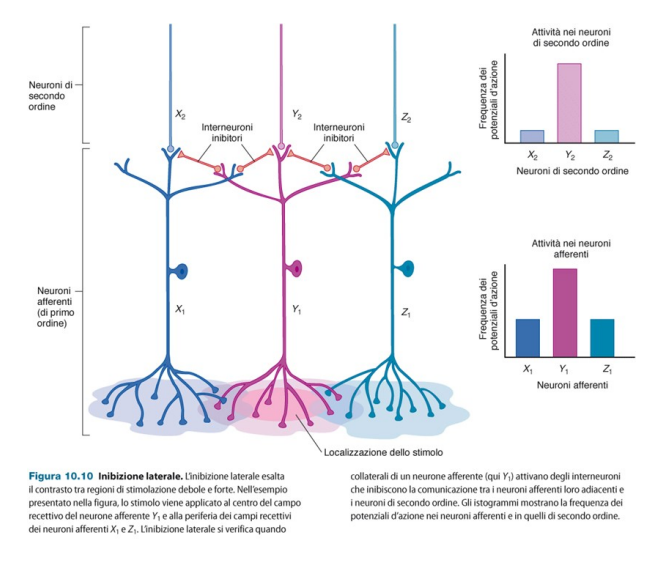
Inibizione laterale.
Per aumentare la precisione di uno stimolo, il neurone più stimolato va ad attivare dei neuroni inibitori. Ciò accade nel caso ci sia una sovrapposizione dei campi recettivi di diversi neuroni.
Codifica dell'intensità dello stimolo
Frequenza dei potenziali d'azione (codifica di frequenza)
Può essere codificata in base a due criteri:
Uno stimolo più intenso può avere un potenziale graduato maggiore. L'ampiezza del potenziale graduato è maggiore (sarà maggiore la frequenza di potenziali d'azione).
Numero di recettori attivi (codifica di popolazione)
Uno stimolo intenso attiverà più recettori.
I recettori sensoriali si distinguono per la soglia d'attivazione. Uno stimolo di bassa intensità attiva quelli a bassa soglia. Uno stimolo a alta intensità attiverà anche quelli con soglia maggiore
Durata dello stimolo
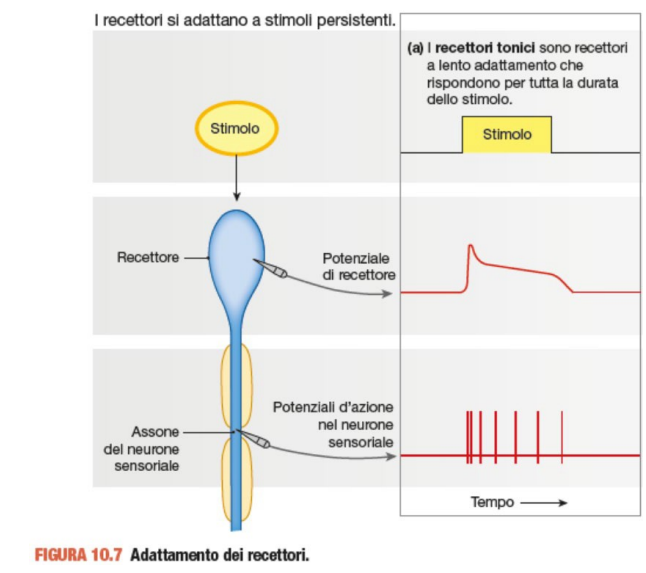
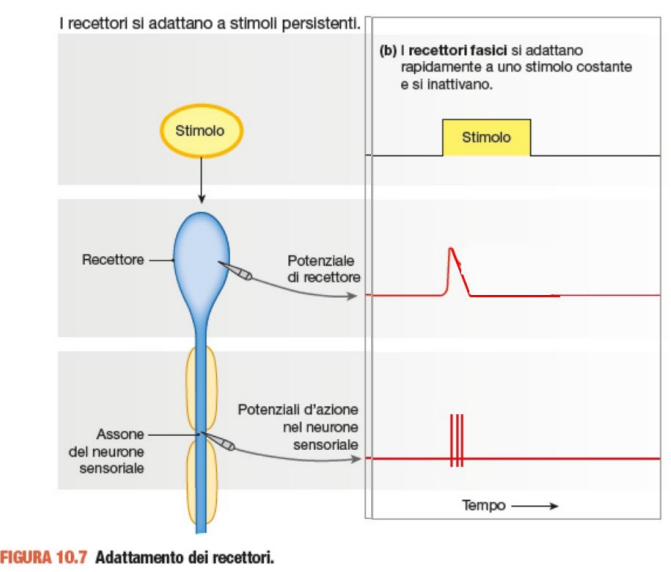
I recettori possono essere distinti in tonici e fasici
I recettori tonici, chiamati anche a lento adattamento. Inizia a scaricare ad alta frequenza. Se lo stimolo persiste il recettore si adatta, ma viene comunicato con una frequenza minore di potenziale d'azione. Si può adattare in maniera parziale o totale (se smette di segnalare lo stato omeostatico).
Un esempio è andare al mare e tuffarsi in acqua. Il recettore ti comunica il cambio di temperatura. Nel tempo però diminuisce la frequenza del potenziale d'azione. L'acqua viene percepita meno fredda, ma restiamo consapevoli di essere in ambiente freddo.
Il recettore fasico segnala solo una novità, un cambiamento. Si adatta immediatamente e niente più potenziali graduati e d'azione. Ricomincia a rispondere solo al variare dello stimolo o il suo terminare.
Sensi somatici
Esterocezione
Propriocezione
Concerne il senso di sè, postura e movimento. Non sempre hanno un correlato coscente. Deriva dall'attività di fusi neuromuscolari, organi tendinei del Golgi e recettori articolari
Enterocezione
Tatto
Ci sono distinte strutture sensoriali.
Cellule di Merkel (recettori non neuronali a lento adattamento, coinvolte nella discriminazione statica delle forme) e corpuscoli di Meissner (mi segnalano il movimento laterale. A rapido adattamento). Sono recettori di superficie, che comunicano cose distinte.
Nel derma ci sono altri recettori
Il corpuscolo di Pacini sono sensibili alle vibrazione. Rispondono a stimoli di contatto a elevata frequenza. Sono fasici. I Corpuscoli di Ruffini sono a lento adattamento, sono sensibili a pressione e distensione cutaenea.
Questi recettori sono utili alla sensibilità epicritica, che ci permette di discriminare tramite un tatto fine.
Poi ci sono terminazioni libere, che riguardano dolore, temperatura e tatto grossolano, definito come sensibilità protopatica.
Le fibre degli stimoli tattili sono a conduzione rapida. La sensazione dolorifica è più lenta (la refrazione è più rapida della coscienza del dolore)
| Cellule di Merkel | Epitelio | Tonico | Discriminazione delle forme |
|---|---|---|---|
| Corpuscoli di Meissner | Epitelio | Fasico | Movimento laterale |
| Corpuscoli di Pacini | Derma | Tonico | Vibrazioni, frequenza elevata |
| Corpuscoli di Ruffini | Derma | Fasico | Distensione |
Recettori termici
I recettori termici sono distinti in base ai range di temperature. Le temperature estreme vengono percepite come dolorosi e non più da termorecettori (nocicettori)
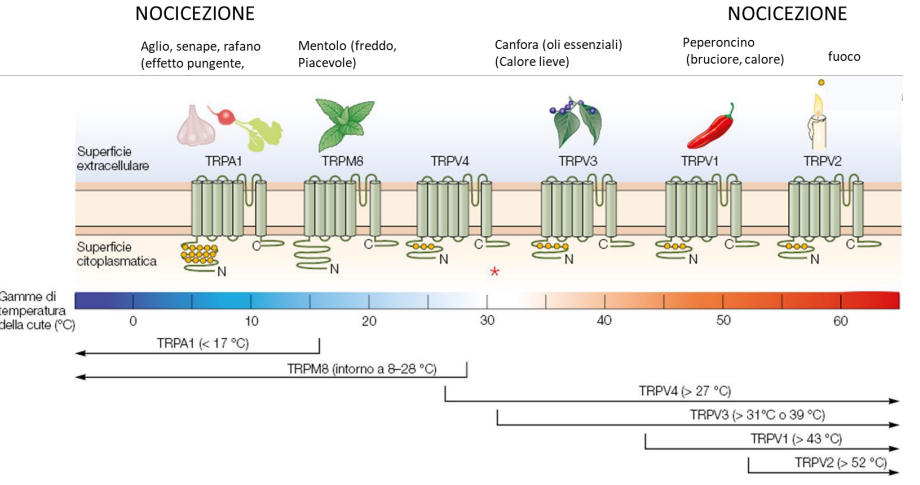
Le fibre afferenti dei nocicettori ascendono con vie distinte e codificano per uno stimolo doloroso e non a uno stimolo di temperatura.
La nocicezione può essere mediata anche da nocicettori meccanici (stimoli dolorifici meccanici) e polimodali (stimoli nocivi di molti tipi, chimici, termici, meccanici). Dolore ritardato, diffuso e duraturo.
Il dolore è mediato da fibre mieliniche distinte (mieliniche nel primo dolore, amieliniche nel secondo)
Iperalgesia, allodinia, dolore riferito e prurito
Durante i processi infiammatori, un nocicettore viene sensibilizzato (iperalgesia); cioè c'è una risposta eccessiva a uno stimolo dolorifico), si abbassa la soglia di risposta allo stimolo.
Si può verificare anche allodinia (cioè c'è una risposta dolorifica ad uno stimolo innocuo).
Il dolore riferito è percepito in aree superficiali del corpo, ma viene generato dalla stimolazione di nocicettori posti a livello di visceri. La corteccia cerebrale non riesce a localizzare in maniera corretta.
Un neurone di 1° ordine dai visceri fanno sinapsi con gli stessi neuroni di 2° ordine che utilizza la superficie corporea, che è più utilizzata.
Prurito
E' mediato da fibre C specifiche, distinte dai nocicettori. Vengono attivati da istamina ...
Potrebbero utilizzare le stesse sottopopolazioni di neuroni coinvolte nei nocicettori. Uno stimolo dolorifico applicato dove si sente prurito inibisce questo stimolo.
Teoria del cancello
Uno stimolo tattile può inibire (parzialmente) uno stimolo dolorifico. La fibra mieilinica nocicettiva fa sinapsi con il suo neurone di 2° ordine. La fibra del recettore tattile fa sinapsi con un neurone inibitore, che può inibire il neurone di 2° ordine, rendendo lo stimolo nocicettivo meno intenso.
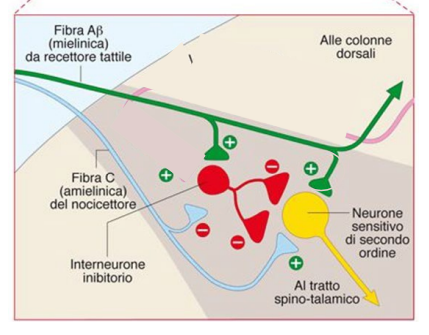
Trasmissione dello stimolo
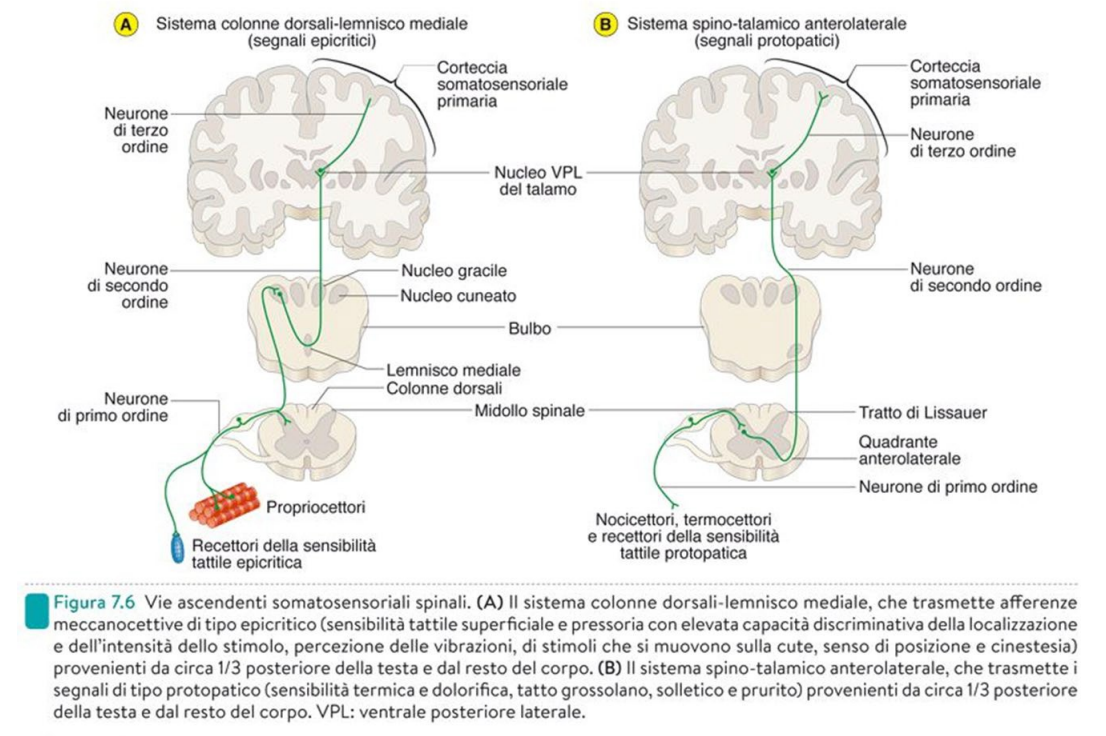
La sinapsi tra neurone 1° e 2° avviene nel Bulbo, non nel midollo. Il neurone di 2° ordine ascende controlateralmente. Sensibilità del lato destro arriva alla corteccia sinistra.
Per i segnali protopatici lo switch avviene a livello del Midollo.
Corteccia somatosensoriale
Ciascuna area e via sensitiva ha una regione nella corteccia che corrisponde al campo sensoriale. L'area della corteccia dedicata ad ogni parte è relazionata alla finezza di discriminazione (Ci sono più neuroni di 3° ordine).
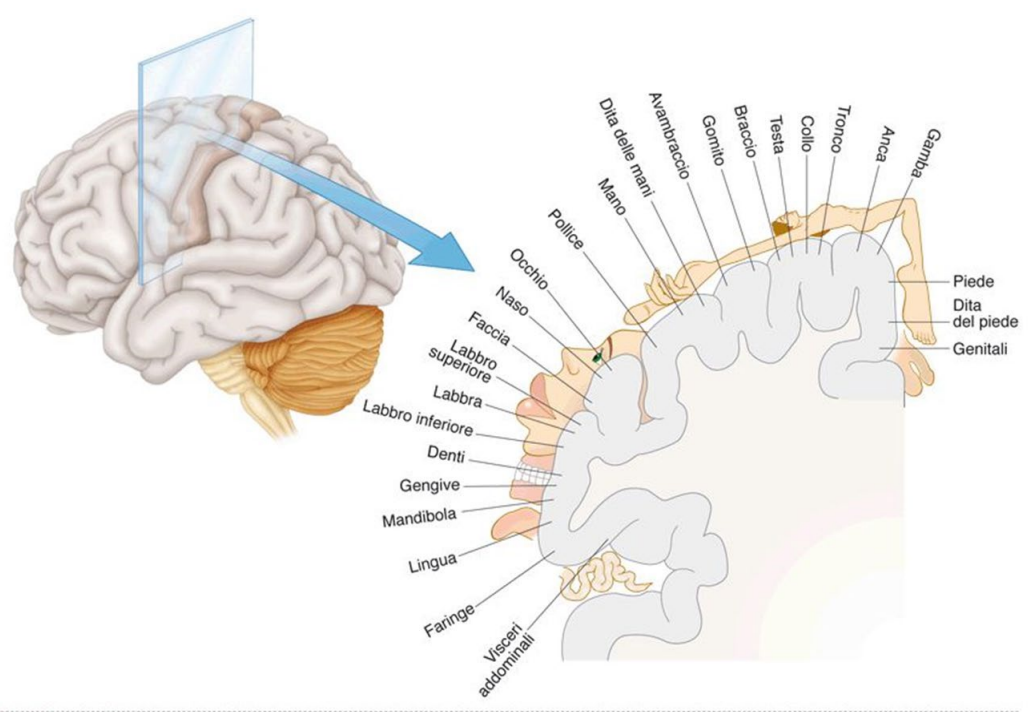
L'olfatto
E' un senso molto antico, coinvolto nella nocicezione. E' l'unico senso associato ad emozione e ricordi. (?)
E' mediato da neuroni bipolari Appr. che si trovano nell'epitelio olfattivo (cavità nasale). Qui abbiamo anche cellule di sostegno e cellule staminali. Infatti i neuroni sono l'unico esempio di cellule nervose che possono rigenerarsi nell'essere umano. Si cambiano ogni due mesi.
Ogni neurone ha un singolo dendrite ed un assone che si estende nel bulbo olfattivo. Ogni neurone olfattivo ha un unico tipo di recettore. Un unico recettore può riconoscere odoranti diversi, un singolo odorante può essere riconosciuto da più recettori.
Trasduzione dei segnali olfattivi
Tutti i recettori non sono disposti in ordine nell'epitelio olfattivo, ma sono in ordine sparso. Li ordina il fatto che recettori dello stesso tipo proiettano nella stessa area del bulbo olfattivo. Proiettano in livelli che chiamiamo glomeoli.
Ciò permette di discriminare meglio tra più odori. Ci permette di ricordare odori anche per anni.
E' l'unica via sensoriale che non passa per il talamo. Alcune vie olfattive dal bulbo giungono all'amigdala e all'ippocampo, componenti del sistema limbico (coinvolti in emozioni e memoria).
C'è un forte adattamento cellulare agli stimoli olfattivi. Dopo pochi minuti però si riesce a resettare.
L'abituazione centrale invece riguarda la plasticità sinaptica, e serve più tempo per riabituarlo.
Il gusto
I recettori del gusto si trovano nel cavo orale, in strutture che sono i xx gustativi.
Abbiamo cellule di supporto e cellule sensoriali, che possono distinguere tra vari sapori.
Possono legare con molecole dette gustanti. Ci sono dolce, salato, amaro, acido, umami
Sodio esogeno stimola i recettori del salato, entra nella cellula e la depolarizza. I recettori dell'acido xx
Dolce, amaro e umami condividono la stessa cascata. Sono tutti mediati dalla gustoducina, che depolarizza e fa rilasciare i neurotrasmettitori.
Dolce -> saccarosio
Amaro -> Composti contenenti azoto (spesso coinvolti nella nociocezione)
Umami -> Esaltatori del sapore, costituiti da amminoacidi, soprattutto glutammato.
Gusto e olfatto contribuiscono alla costruzione del sapore. E' una metasensazione, data dalla combinazione dei composti aromatici del cibo che raggiungono la cavità retronasale. Si aggiunge anche il ruolo dei recettori somatosensoriale della cavità del cibo (consistenza del cibo).
L'udito
Percezione di onde sonore, ovvero onde pressorie che alternano zone di aria compressa e rarefatta.
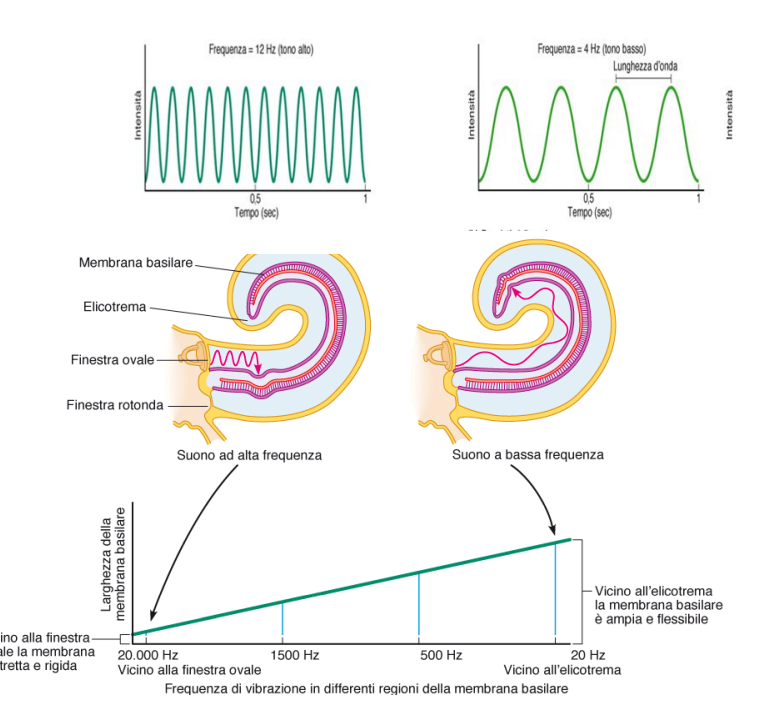
E' misurata in cicli al secondo o Hz. e dipende dalla lunghezza dell'onda. Percepiamo i suoni come bassi o acuti in base alla frequenza.
La potenza o intensità si misura in Decibel. Oltre i 120 dB si percepisce dolore. Una conversazione normale è attorno ai 60 dB.
Nell'orecchio interno troviamo la coclea, struttura ossea a forma di spirale la cui estremità è chiamata elicotrema. La sezione trasversale ci mostra che i compartimenti sono separati da due membrane (vestibolare e basilare, dall'alto verso il basso). La basilare è coinvolta nella trasduzinoe del segnale.
I tre compartimenti sono chiamati scala vestibolare, media e timpanica. All'interno della coclea scorre un liquido.
Scala vestibolare e timpanica il liquido è la perilinfa, nella scala media troviamo l'endolinfa.
La scala è connessa alla finestra ovale dalla staffa.
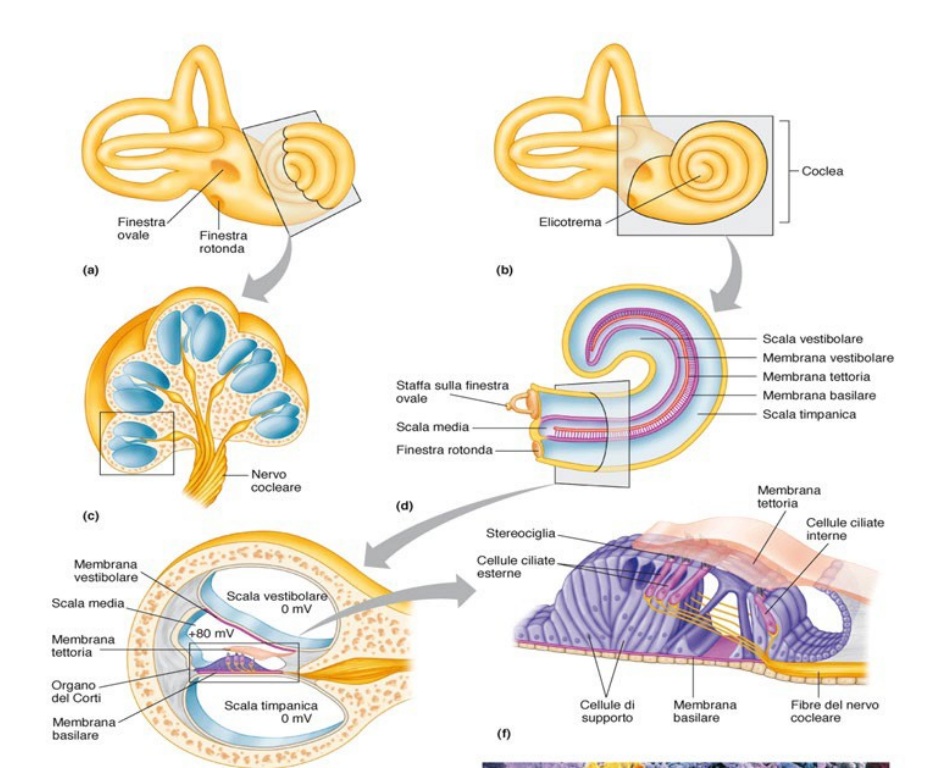
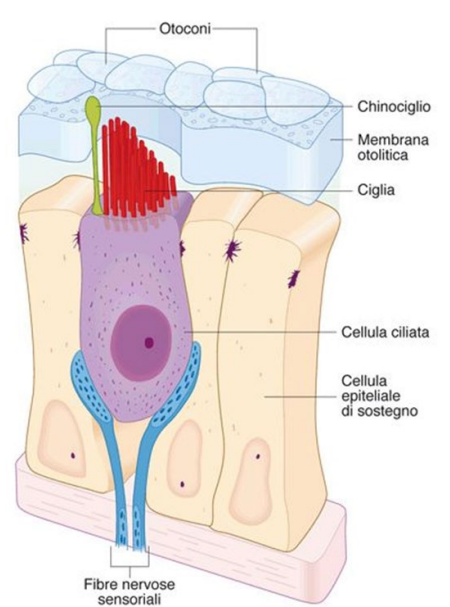
L'organo del Corti appoggia sulla membrana basilare. All'interno troviamo una serie di cellule. Ci sono diverse cellule di sostegno, con poi cellule ciliate, che sono le cellule recettorie. Sono dotate di stereociglia. Sono disposte in ordine in base all'altezza delle stereociglia.
Sono inserite nella membrana tectoria.
Il suono passa da timpano, che fa oscillare incudine e martello, che riverberano sulla staffa. Servono ad amplificare il segnale. La staffa genera onde lungo la coclea. La pressione generata viene scaricata dalla finestra rotonda (apertura membranosa).
Alla base della traduzione del suono, ci sono queste onde che spingono sulla membrana e la deformano. Quando la membrana basilare si deforma, va a piegare le stereociglia.
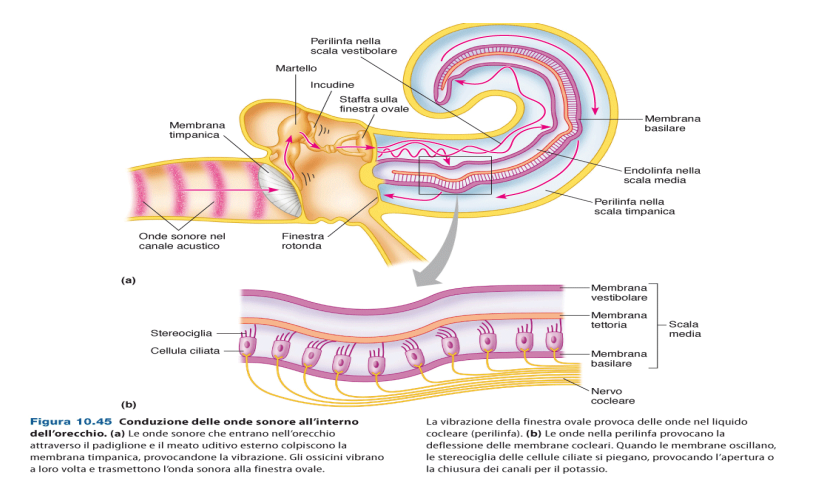
All'apice delle stereociglia troviamo dei canali meccano-sensibili. Le ciglia sono legate da ponti proteici, che collegano anche canali meccano-sensibili, chiamati Canale MET.
In assenza di stimoli, questi canali sono sempre parzialmente aperti. Tengono la cellula sempre parzialmente depolarizzata. L'endolinfa è ricca in potassio. Il potenziale è di -60 mV. Ciò permette una rapida risposta agli stimoli.
Quando si piegano le stereociglia in direzione del stereociglio più alto. I ponti protici entrano in azione e fanno aprire i canali meccano-sensibili. Entra potassio e la cellula depolarizza. Quando si piegano in direzione opposta, verso lo stereociglio più basso i canali si chiudono e la cellula iperpolarizza.
Verso l'alto -> Depolarizza
Verso il basso -> Iperpolarizza
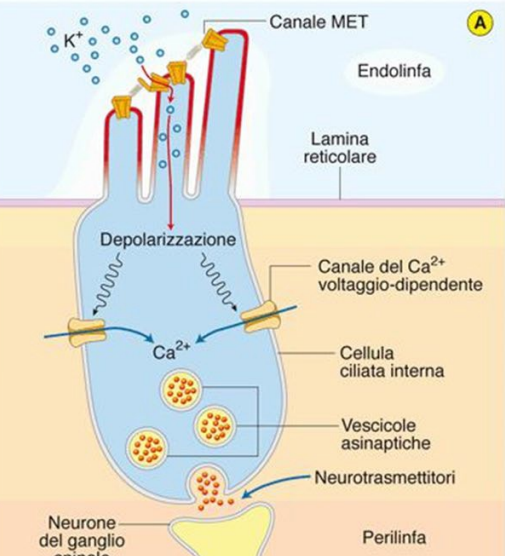
Lo scorrimento relativo della membrana tectoria è ciò che da la direzione alle stereociglia. La membrana basilare è ciò che da il movimento, la membrana tettoria è più stabile e scorre di conseguenza.
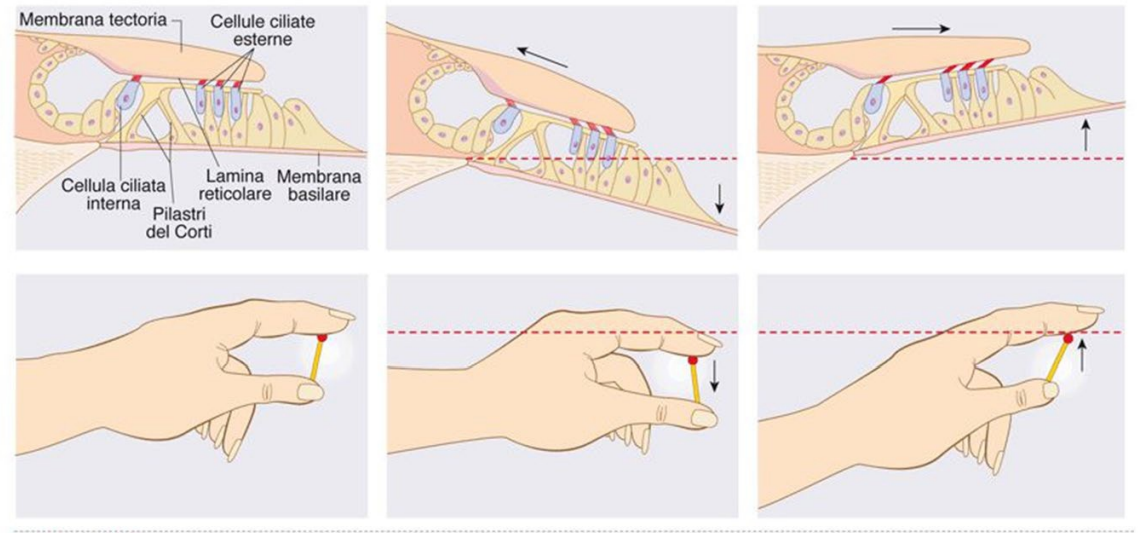
Codificazione di intensità e tonalità del suono
L'intensità si codifica facilmente, siccome un suono forte causa un reclutamento maggiore di cellule ciliate.
La frequenza dipende da una proprietà della membrana basilare. E' più rigida in prossimità della finestra ovale, ed è più flessibile in prossimità dell'elicotrema.
I suoni ad alta frequenza stimoleranno in prossimità della finestra ovale, mentre quelli a bassa frequenza stimoleranno la porzione vicina all'elicotrema.
Equilibrio
L'apparato vestibolare è pure collocato nell'orecchio interno. E' responsabile del senso dell'equilibrio, siccome rileva i movimenti della testa.
Non è coinvolto solo questo apparato. Abbiamo continuamente dei feedback dal sistema somato-sensoriale che contribuisce sulla percezione del corpo.
L'apparato vestibolare rileva i movimenti angolari e di accelerazione, assieme alla pos della testa rispetto al corpo.
Ci sono canali semicircolari, otriculo e sacculo.
I canali semicircolari rilevano i movimenti angolari della testa e sono orientati sui tre assi. Otriculo e sacculo rilevano i movimenti lineari.
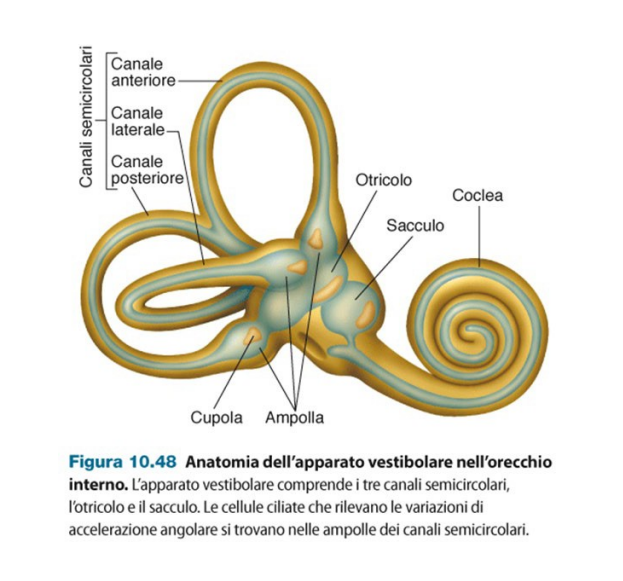
| Canale anteriore | Movimento verticale-saggittale (Alto-Basso) |
| Canale posteriore | Movimento verticale-frontale (Orecchio-spalla) |
| Canale laterale | Movimento laterale (testa per lato) |
| Otriculo | |
| Sacculo | |
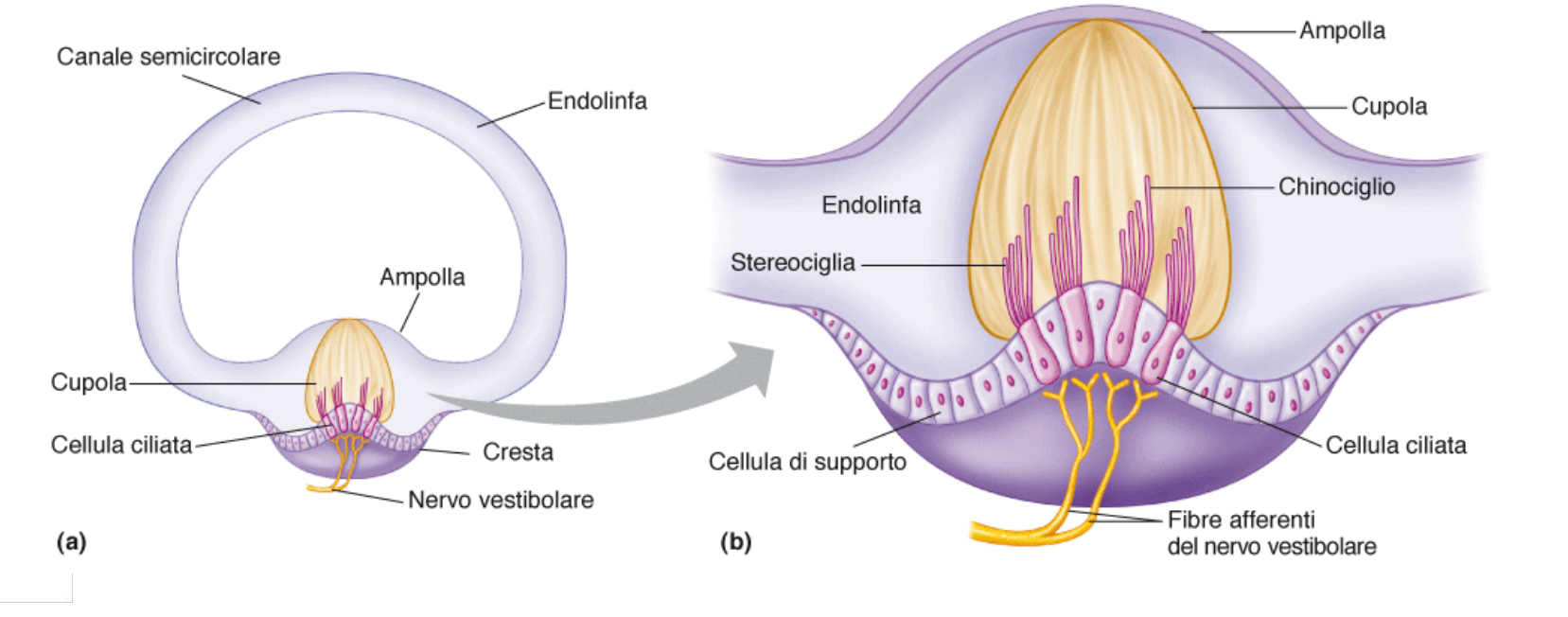
| L'endolinfa scorre all'interno di questi canali. Alla base dei canali troviamo l'ampolla; costituita da cresta e cupola. | |
| Anche qui troviamo cellule di supporto e cellule ciliate. Qui pure troviamo stereociglia poste in oridne di grandezza. Qui però si il ciglio più alto è chiamato chinociglio, ed è dotato di microtubuli. | |
| Quando ruotiamo la testa l'endolinfa si muove per inerzia in senso opposto. Ciò fa ruotare la cupola e girare l'ampolla. Si muovono quindi le ciglia, e la cellula deporalizza se si muovono verso lo stereociglio e iperpolarizza al contrario | |
| Partono canali afferenti. |
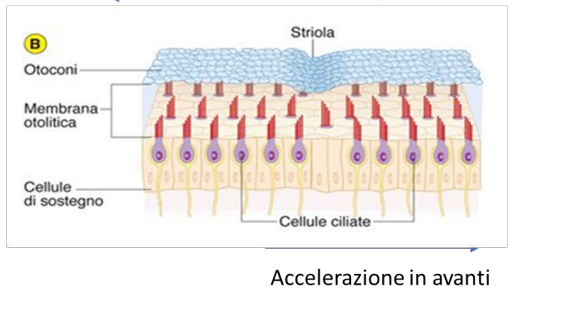
Otriculo e sacculo hanno lo stesso sistema. Qui sono inserite in na sorta di membrana gelatinosa, con appoggiati sopra otoliti, cristalli di carbonati di calcio. Quando c'è un movimento gli otoconi per inerzia andranno dalla direzione opposta; faranno piegare le stereociglia in quella direzione.
Movimento verso il chinociglio -> depolarizzazione.
Essendo che devono rilevare movimenti lineari (accelerazioni)
Per poter percepire lo spostamento in entrambe le direzioni, le cellule sono poste in maniera speculare.
Vista
I nostri recettori riescono a trasdurre onde elettromagnetiche nella lunghezza d'onda della luce visibile (350-750nm).
Ciò avviene grazie alle proprietà della luce di essere riflessa (parte della luce non viene assorbita e viene invece riflessa, noi percepiamo quelle lunghezze d'onda) e rifratta (la luce cambia direzione in base alle proprietà delle superfici che incontra).
La rifrazione avviene all'interno dell'occhio, grazie alla presenza del cristallino, struttura biconvessa formata da cellule elastiche (che perdono nucleo e organelli), formando una lente che va a concentrare la luce in un punto, detto fuoco.
Alla base del bulbo oculare troviamo la retina, dove si collocano anche i fotorecettori. Per una visione ottimale, il fuoco deve cadere a livello della retina.
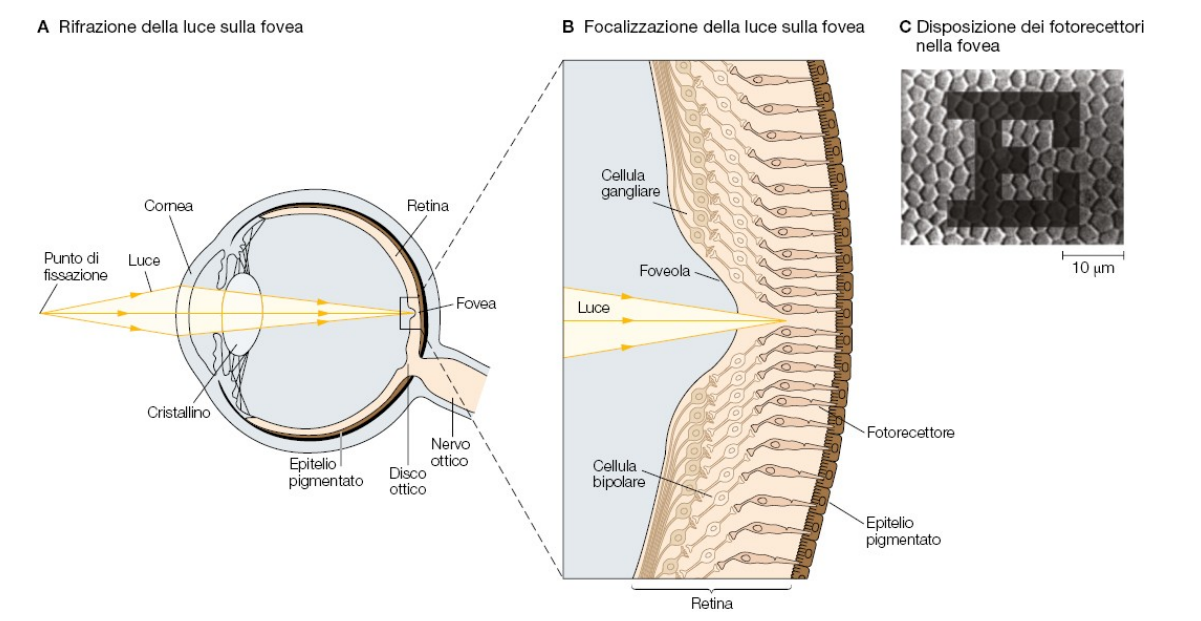
L'immagine viene proiettata sulla retina al contrario, è la corteccia a capovolgerla. La fovea è l'area di massima acuità visiva, dove abbiamo il maggior numero di recettori.
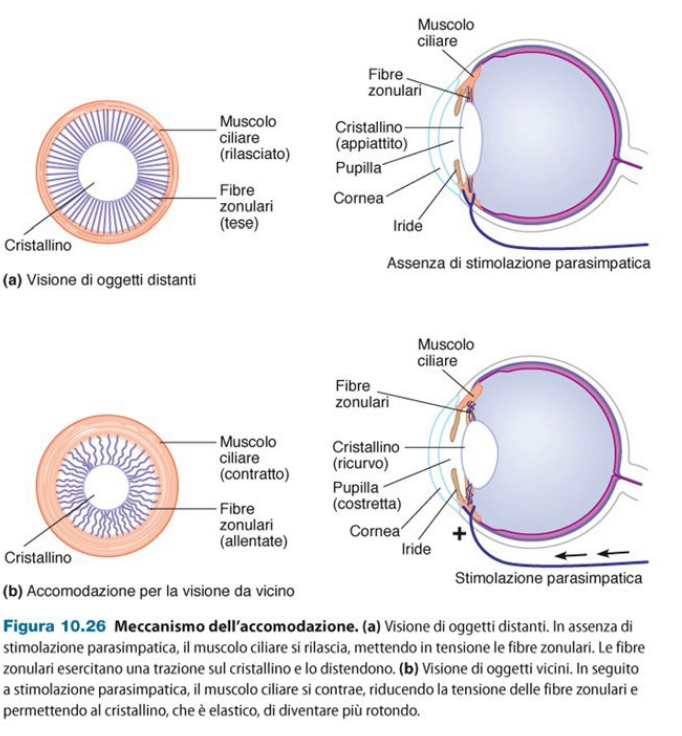
Fino ad una distanza di 6m i raggi luminosi arrivano paralleli al cristallino. Quando ci avviciniamo ad un oggetto, i raggi luminosi diventano divergenti. Perciò il cristallino deve accomodare (diventare più ricurvo) in maniera che il fuoco cada sempre sulla retina. E' un riflesso mediato dal sistema parasimpatico. Il cristallino è legato al muscolo ciliare da fibre zoonulari. Normalmente queste fibre lo tengono disteso, all'avvicinarsi di un oggetto lo contraggono e si ricurva
Fototrasduzione
Come avviene la fototrasduzione?
Avviene a carico dei fotorecettori (che sono Coni e Bastoncelli), che comunicano con le cellule bipolari, che comunicano a loro volta con le cellule gangliari, dove si forma il nervo ottico.
C'è un punto dove emerge il nervo ottico, qui non vediamo ed è un punto cieco.
Nel segmento interno sono collocati i xx
Bastoncelli sono costituiti da numerose invaginazioni che formano una serie di dischi. Nel segmento esterno troviamo i fotopigmenti, che contengono una molecola di retinale e una di opsina. I bastoncelli contengono solo la rodopsina, i coni invece possono contenere diversi tipi di opsine, sensibili a diverse lunghezze d'onda.
I bastoncelli ci permettono di vedere al buio, siccome sono più sensibili, ma non ci consentono di distinguere i colori (monocromatico).
I coni invece sono meno sensibili.
La visione, oltre al colore, si basa sui contrasti, aumenti e diminuzioni di quantità di luce disponibile. Entrambe le informazioni vengono inviate alla corteccia.
xx sono in connessione con due tipi di cellule bipolari, con due tipi di recettori diversi.
Ampa/Kainato
mGluR6
I recettori sono in sinapsi con due cellule bipolari ON/OFF, con recettori diversi per lo stesso neurotrasmettitore (glutammato).
Quando c'è poca luce, il recettore è sempre depolarizzato, grazie all'azione del GMPciclico, che mantiene aperti i canali del sodio. E' quindi costantemente depolarizzata e con potenziale graduato viene trasdotto un segnale per cui viene rilasciato costantemente glutammato (neurotrasmettitore).
Quando c'è poca luce, e quindi rilascio di glutammato più intenso, le cellule bipolari ON si iperpolarizzano (perchè è una sinapsi inibitoria).
Nelle cellule bipolari OFF, in caso di poca luce/diminuzione, si depolarizzano e segnalano una diminuzione della luce.
Quando arriva luce, viene assorbita dal fotopigmento.
Il retinale diventa retinale trans associandosi alla rodopsina, cambia conformazione. Fa attivare l'opsina che fa attivare la trasducina. Questa attiva una fosfodiesterasi che degrada il GMPciclico. Si chiudono i canali del sodio.
Il fotorecettore quindi tende a iperpolarizzarsi. Viene rilasciato meno glutammato. Le cellule ON quindi non vengono più inibite, i canali cationici si aprono e le cellule bipolari ON depolarizzano. Segnalano alla corteccia che la luce è aumentata.
Meno glutammato causano una minore eccitazione dei Bipolari OFF, che iperpolarizzano.
I bastoncelli possono saturare, se tutto il retinale cambia di conformazione. (Poi deve dissociarsi dalla rodopsina) e riconvertirsi a retinale cis. Quando cambia velocemente la quantità di luce disponibile, c'è uno scarto nel riadattamento ai livelli di luce perchè il retinale deve assumere la conformazione corretta. Alla luce i bastoncelli si "sbiancano" completamente e intervengono i coni.
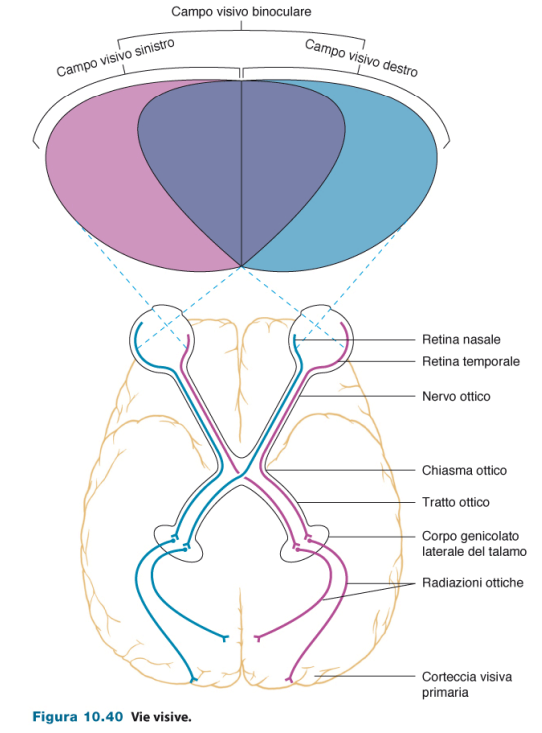
Campo visivo
Il campo visivo destro è costituito da retina temporale sinistra e retina nasale destra. Il contrario per il campo visivo sinistro (retina temporale destra e retina nasale sinistra). Il campo visivo binoculare è percepito da entrambi gli occhi e ci permette quindi di percepire la profondità.
C'è un punto in cui si incrociano le fibre provenienti dalle retine nasali. Gli assoni decussano nel chiasma ottico. In questo modo tutto il campo visivo destro arriva all'emisfero sinistro, e tutto il campo visivo sinistro arriva all'emisfero destro.