I muscoli scheletrici sono generalmente collegati a due ossa.
Non tutti i muscoli sono collegati ad ossa (ad esempio i muscoli legati alla cute o a cartilagini). Ci sono anche muscoli connessi ad altri muscoli (sfintere anale). Tra i muscoli e le ossa si trovano i tendini, che servono a trasferire la forza muscolare ai tendini.
Struttura muscolare
Epimisio: cartilagine che copre il muscolo, che però poi diventa tendine quando collegato all'osso.
Perimisio: Tessuto connettivale
Fascicolo: "pacchetto" di fibre muscolari
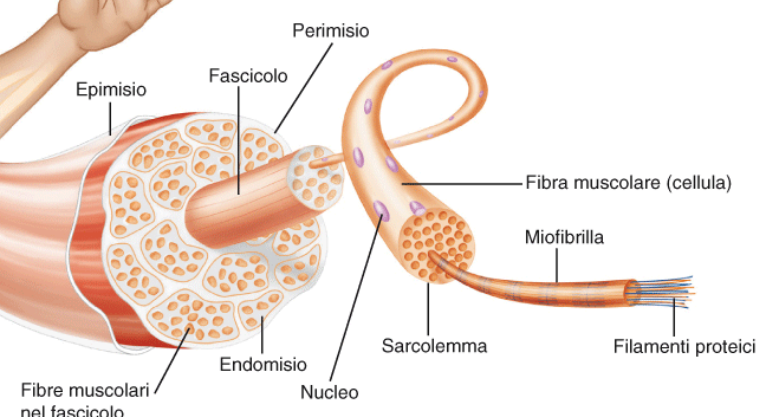
Ogni fascicolo è formato da Fibre muscolari, insieme di cellule (fuse tra loro) costituita da tanti nuclei. Ogni fibra muscolare è lunga come il muscolo. Decorrono parallelamente e diagonalmente rispetto al muscolo. Ciascun fascicolo contiene da centinaia a migliaia di fibrocellule.
Ciascuna fibriocellula è circondata da endomisio. A livello microscopico, dentro ad ogni fibra muscolare, decorrono le miofibrille, costituite da un insieme di filamenti proteici, che costituiscono la vera parte contrattile del muscolo.
Sarcolemma è la membrana plasmatica della fibra. Il citoplasma è chiamato sarcoplasma.
Struttura cellulare
Reticolo sarcoplasmatica, circonda ciascun miofibrilla ed è in associazione ai Tubuli trasversi (Tubuli T) che si connettono ad altre strutture, tra queste ci sono le cisterne laterali, che sono grandi "cisterne" di ioni calcio.
A loro volta sono connessi al sarcolemma e penetrano all'interno della cellula.
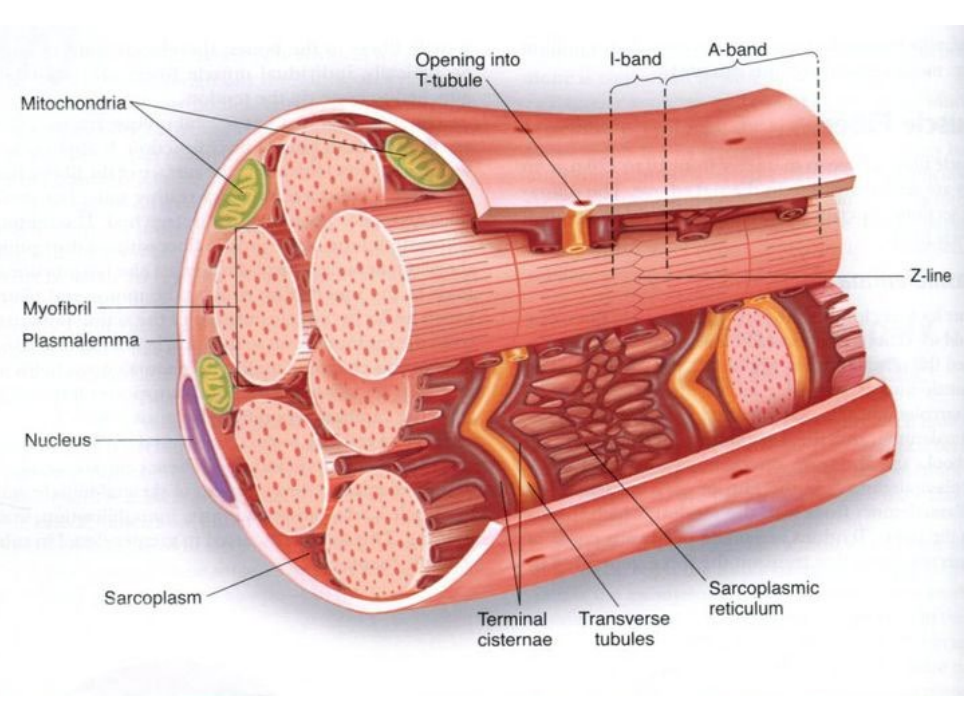
Triade: due cisterne e tubulo trasverso.
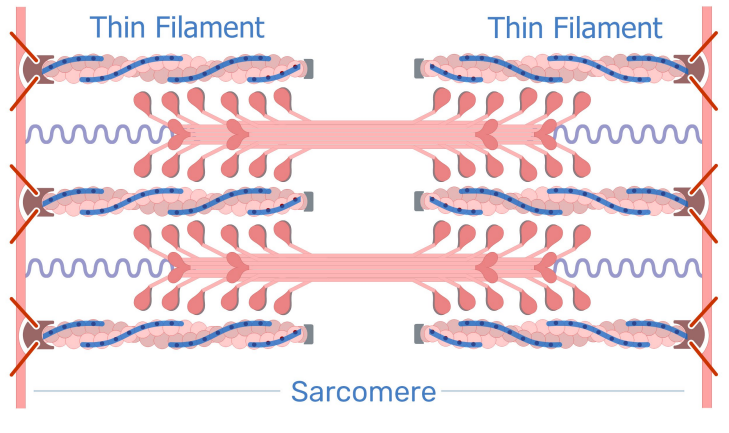
Struttura molecolare
I componenti di filamenti sottili sono monomeri di actina, mentre i filamenti spessi sono dati dalla miosina.
Da questi filamenti origina l'unità funzionale fondamentale del muscolo, chiamata sarcomero. E' un insieme ordinato di filamenti sottili intercalato da filamenti spessi. La contrazione muscolare è l'accorciamento del sarcomero. Il filamento sottile scorre su quello spesso, facendo sparire gli spazi tra i filamenti sottili.
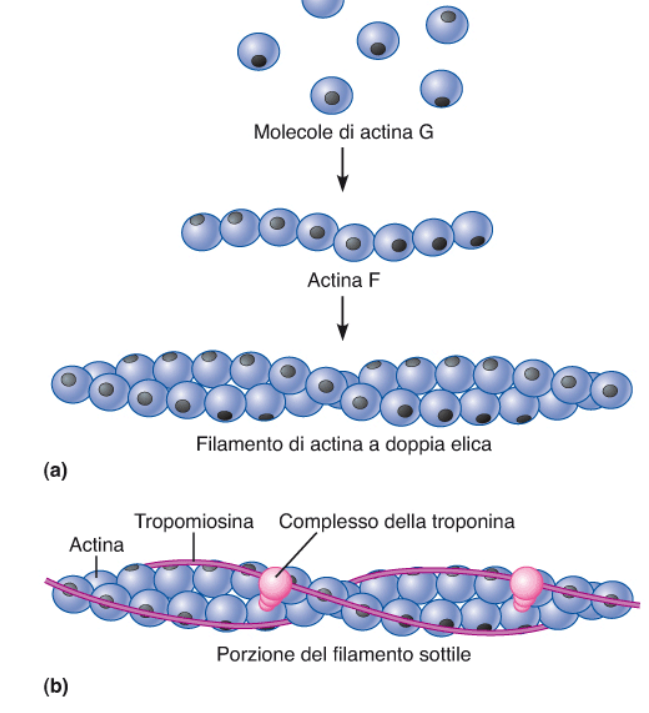
Il filamento di actina è formato da monomeri di actina, che creano dei polimeri, organizzati in filamenti a doppia elica che espongono i siti di legame per la miosina. La tropomiosina è un filamento regolativo che occupa i siti di legame della miosina quando il muscolo è a riposo. La presenza di ioni calcio (che si legano alla tropomiosina) innesca la liberazione di questi siti di legame alla miosina.
I filamenti spessi sono costituiti da centinaia di molecole di miosina, ciascuna molecola è un dimero costituito da due code e due coppie di teste.
La testa è la “parte che lavora” della molecola di miosina, poiché essa è la parte che genera attivamente la forza meccanica sviluppata dal muscolo. Ciascuna testa possiede due siti fondamentali per la generazione della forza contrattile: un sito di legame per l’actina, in grado di legarsi ai monomeri di actina dei filamenti sottili, e un sito ATPasico, che possiede attività enzimatica e idrolizza l’ATP.
Generazione della forza muscolare
Le actine scorrono sulle miosine. Il risultato è che con la contrazione il sarcomero si accorcia. Diminuisce lo spazio della zona H, si avvicinano i lati del sarcomero.
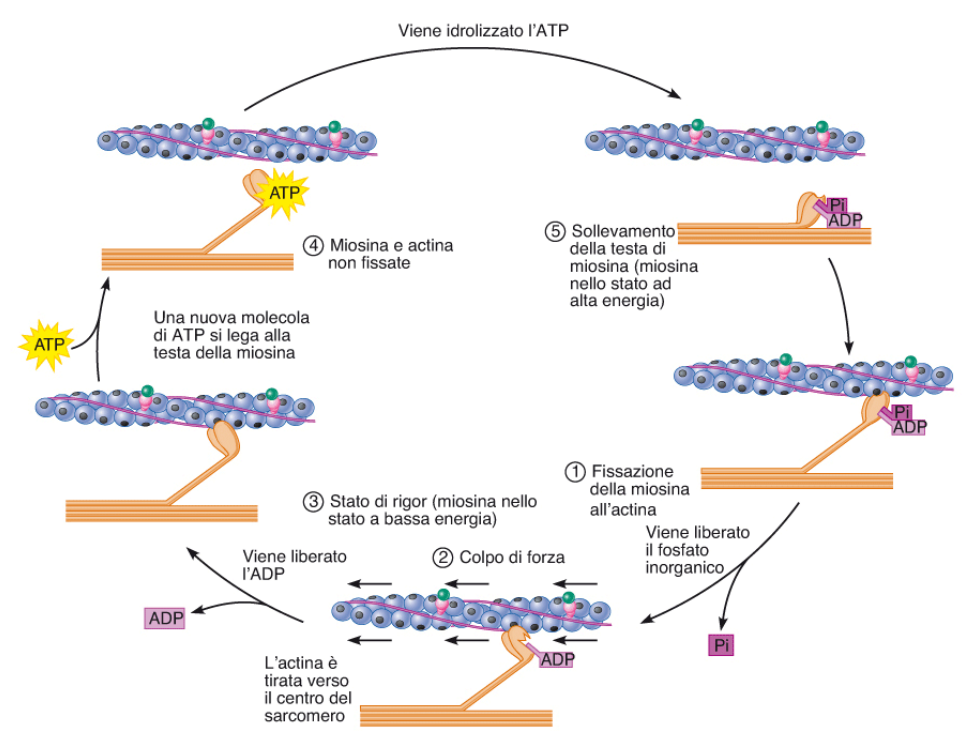
La contrazione inizia con l'aggancio tra miosina e actina.
Per lo scorrimento è necessario il ciclo dei ponti trasversali.
1 - Il calcio si lega alla tropomiosina, si sposta e libera i siti della actina. Actina e miosina si legano. Nel sito della miosina che lega ATP in questo momento c'è ADP+P. In questo momento però viene liberato il fosfato inorganico P.
2 - La liberazione del fosfato inorganico genera un cambiamento della conformazione della testa della miosina (piegata), che si piega e fa scorrere il filamento di actina. Questo scorrimento è chiamato "colpo di forza". Questo stato è quello a minore energia.
3 - Viene liberato l'ADP
4 - Arriva ATP, che is lega alla testa della Miosina, che torna alla conformazione iniziale e la stacca dalla actina.
5 - Avviene l'idrolisi dell'ATP in ADP+P, si torna al punto di partenza. (Miosina ad alta energia).
Come si attivano i muscoli
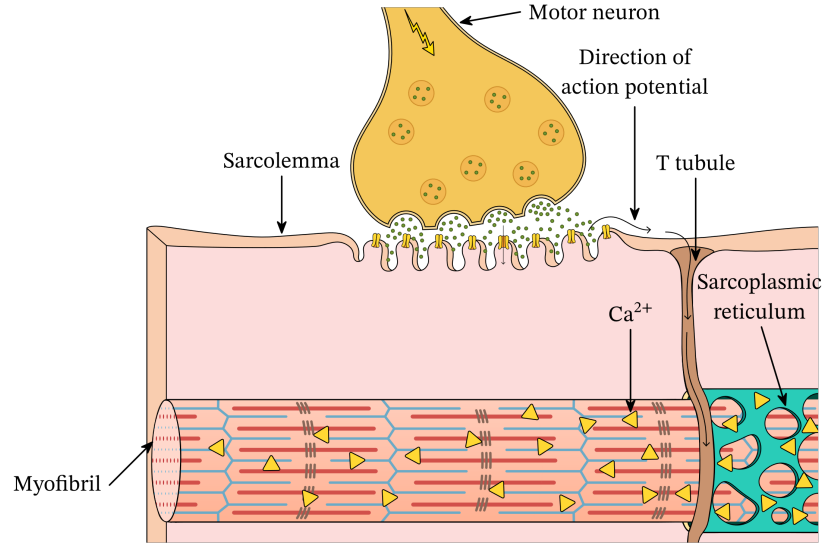
Giunzione neuromuscolare è il punto di giunzione tra motoneurone e fibra muscolare.
Vescicole sinaptiche, viene liberato il neurotrasmettitore in questo caso acetilcolina.
Tubulo T: porta la depolarizzazione che avviene in seguito all'arrivo dell'acetilcolina sulla membrana postsinaptica all'interno della fibra muscolare.
Motoneurone nel terminale asssonico. Nella fessura sinaptica c'è la placca motrice, con molti ricevitori per l'acetilcolina. Si genera una depolarizzazione (potenziale d'azione) che si propaga per i tubuli T. Questi scendono lungo la fibra muscolare e collegano alle cisterne del reticolo sarcoplasmatico.
C'è quindi una vicinanza tra il tubulo T e le cisterne del reticolo sarcoplasmatico che contengono calcio. Questo potenziale determina il rilascio di calcio, che si libera nel sarcoplasma e può raggiungere l'apparato contrattile, cioè i filamenti di actina, sbloccando i suoi siti attivi attraverso il legame con la tropolina (allontanando la tropomiosina). Qui parte il ciclo dei ponti trasversali.
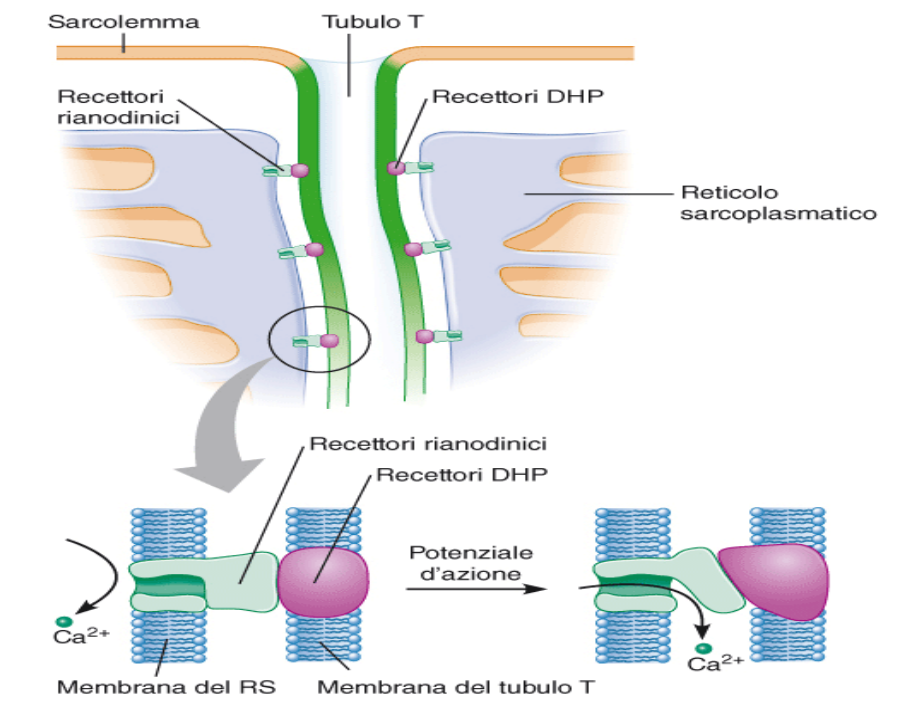
Quando il potenziale d'azione scorre lungo il tubulo T, trova lungo questa membrana dei recettori DHP collegati a recettori rianodinici, posti sulla membrana del sarcolemma (reticolo sarcoplasmatico). Le due membrane hanno recettori connessi.
Quando arriva il potenziale d'azione, il recettore DHP (che è essenzialmente un recettore di voltaggio) cambia di conformazione che a sua volta cambia il recettore rianodinico, che cambia a sua volta conformazione e apre il canale per il calcio. Il risultato è un rapido aumento di calcio nel citosol.
Perchè nel citosol è bassa la concentrazione di calcio, mentre nel reticolo sarcoplasmatico è alta? Perchè sulla membrana del reticolo sarcoplasmatico ci sono pompe attive che pompano continuamente calcio verso il reticolo sarcoplasmatico (Altro motivo di consumo di ATP nei muscoli)
Grazie all’azione di tali pompe, il RS è in grado di accumulare calcio contro gradiente di concentrazione e di svolgere quindi la funzione di sito di immagazzinamento per il calcio.
Come si disattivano
Il calcio fuoriesce RS e si diffonde nel citosol. Quest'uscita deve essere "esplosiva", per poter dare il massimo della forza in un lasso di tempo molto basso.
Sono presenti altri recettori sul RS che sentono il calcio e aprono altri canali calcio (effetto a cascata). Continua il rilascio calcio-dipendente. A livelli molto elevati questi canali si chiudono.
Se cessa l'impulso nervoso, ci troviamo con una elevata concentrazione di calcio nel citosol. Ma i recettori rianodinici sono chiusi, l'elevata concentrazione di calcio che quindi chiude il rilascio calcio-dipendente. Rimane l'attività delle pompe che pompa calcio dal citosol al reticolo.
Quindi la contrazione cessa, perchè crolla la concentrazione del calcio nel citosol.
La tropomiosina torna alla posizione corretta, rendendo indisponibile la miosina.
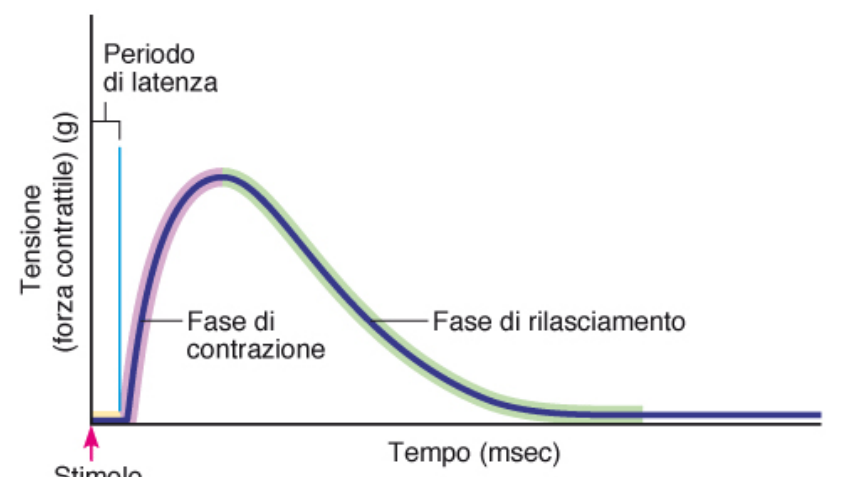
Scossa singola
Il periodo di latenza è quel lasso di tempo in cui il potenziale d'azione si diffonde, ma ancora non viene generata forza. la fase di rilasciamento è la fase più lunga, in cui si ripristina la condizione di partenza.
Contrazione isometrica e isotonica
La forza generata da un muscolo dipende da due fattori:
- la forza sviluppata dalle singole fibre muscolari
- il numero di fibre muscolari che si contraggono
La modulazione della forza di un muscolo dipende dalla quantità di fibre coinvolte.
xxx
Frequenza di stimolazione
A frequenze elevate la velocità con cui il calcio viene liberato dal RS all'interno del citosol supera quella con cui il calcio viene attivamente recuperato dal citosol verso l'interno del RS. Per ciò la tensione del picco aumenta, aumenta quindi la forza fino ad arrivare a un plateu.
C'è quindi una sommazione del calcio, quindi anche la tensione aumenta.
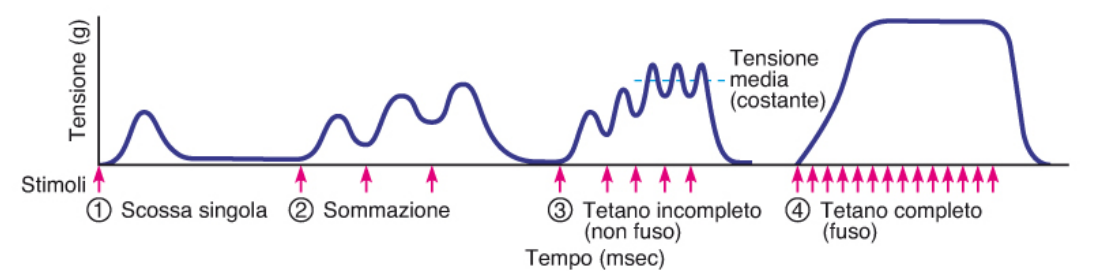
Il tetano rappresenta l'apice, (il nome ha origine patologica), della frequenza delle scariche di stimolazione, dove il muscolo non rilascia più ed ogni fibra è reclutata nel ciclo dei ponti trasversali.
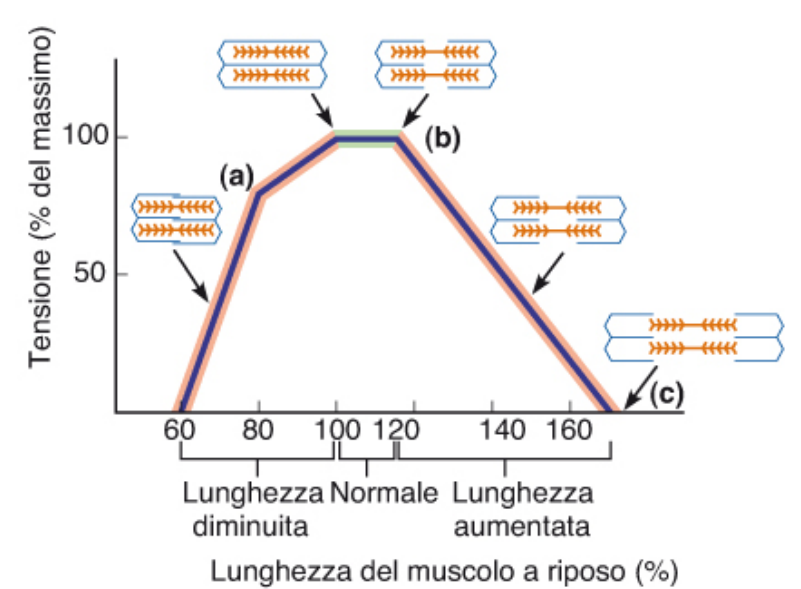
Alla lunghezza (stiramento) del muscolo è anche associata una diversa tensione massima. All'aumentare della lunghezza diminuisce la tensione, siccome c'è minore sovrapposizione tra le unità funzionali.
La lunghezza ottimale (detta normale) è quella in cui il muscolo esprime la forza massima. Alla lunghezza normale avviene la massima interazione tra miosina e actina.
La forza dei muscoli può anche essere modulata tramite la frequenza dei potenziali d'azione. Aumentandoli aumenta la quantità di calcio rilasciata. Aumenta quindi la interazione tra actina e miosina.
Una seconda modalità dipende dal numero delle fibre muscolari coinvolte. La variazione della frequenza dei potenziali d'azione copre solo in piccola parte l'ampio ambito delle forze diverse che il muscolo può sviluppare. Solo una piccola parte delle fibre si muove attivamente, le altre sono passive.
Il reclutamento è il fenomeno per cui il numero di fibre attivate interno a un muscolo aumenta.
Quando una contrazione viene mantenuta per un tempo prolungato, le unità motorie sono attivate in maniera asincrona. Quando una viene attivata, le altre cessano. Così la forza totale è mantenuta a livello costante, evitando che alcune tra le unità motorie attive vengano sovraccaricate.
La velocità di accorciamento è la variazione di lunghezza possibile di un muscolo in funzione del tempo. La velocità dipende anche dal carico che possono supportare.
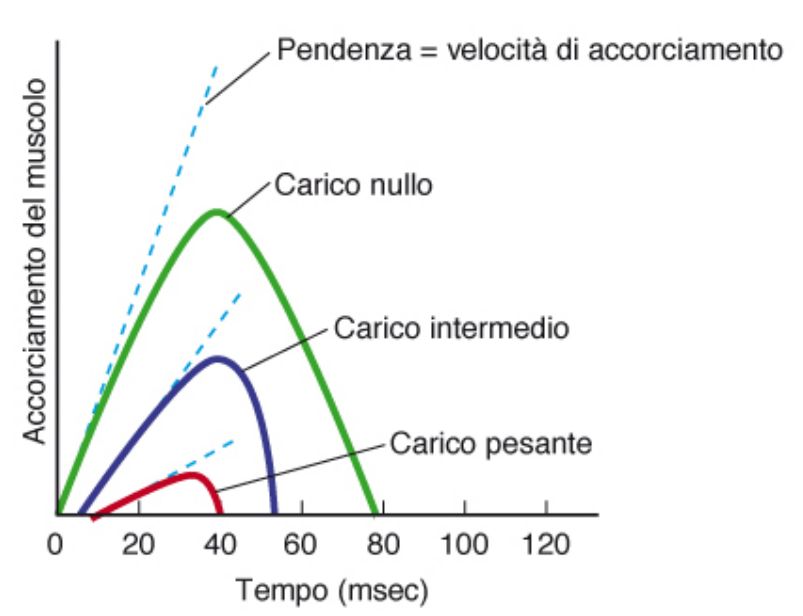
Al crescere del carico la velocità di accorciamento diminuisce, fino a raggiungere un massimo in cui il carico è uguale alla tensione massima del muscolo.
Metabolismo del muscolo scheletrico
Ci sono tre modalità con cui il muscolo può generare ATP:
- Glicolisi anaeroba, che però ha una bassa resa di produzione di ATP (e produzione di lattato).
- In condizioni aerobe, il piruvato non va in lattato ma viene coinvolto nel ciclo di Krebs. Ma è necessario ossigeno
- Creatinfosfato, è una maniera di immagazzinare ATP.
I muscoli scheletrici hanno una scorta di ATP da usare in maniera immediata, per far fronte a situazioni improvvise. Nei primi istanti il muscolo può usare l'ATP immagazzinato. Se il lavoro continua, il muscolo utilizzerà la glicolisi e il ciclo di Krebs. Tuttavia l'apporto di ossigeno può non essere sufficiente. Qui inizia la produzione di acido lattico, causando però una diminuzione della performance del muscolo.
La maggior parte dell'ATP che produciamo durante i primi momenti dell'esercizio proviene dalla glicolisi anaerobica, soprattutto negli esercizi più intensi. La fosforilazione ossidativa è il metodo che contribuisce di più ATP, ma ci mette di più ad "attivarsi".
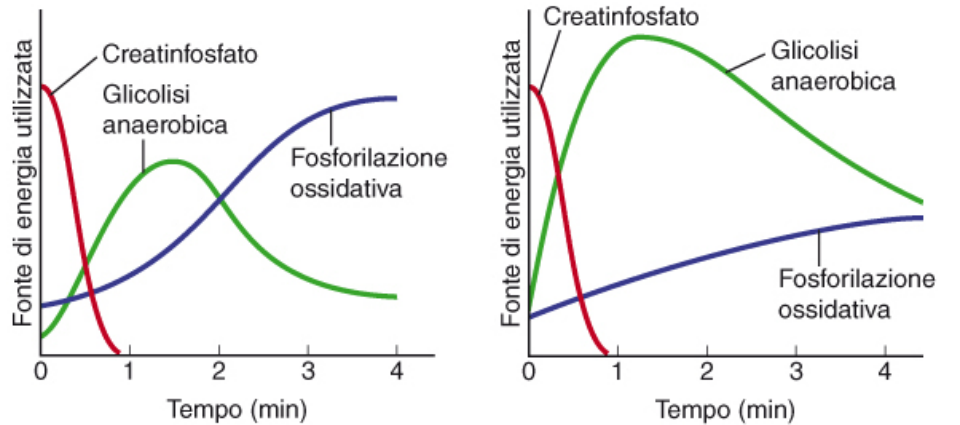
La fosforillazione ossidativa produce circa 36 ATP da una molecola di glucosio. La glicolisi anaerobica ne produce solo 2, assieme allo scarto rappresentato dall'acido lattico.
Fibre lente e veloci
I muscoli dell'occhio sono fibre molto veloci. Nella gamba (gastrocnemio) la proporzione di fibre rapide e lente è circa uguale. Nel soleo della gamba sono contenute fibre più lente.
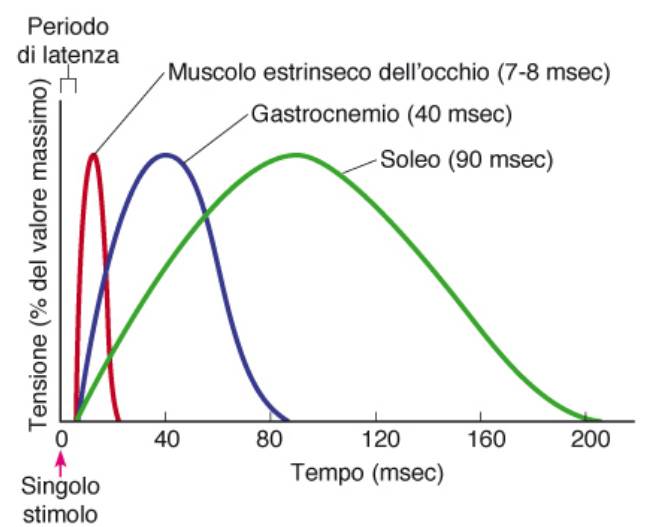
La differenza è data dal tipo di miosina che costituisce i filamenti spessi. C'è una miosina rapida che riesce a idrolizzare l'ATP più velocemente, riesce quindi a contrarre le fibre più rapidamente.
Le fibre glicolitiche o ossidative sono fibre distinte che riescono ad attivare con maggiore efficienza certi pathway metabolici, hanno quindi un diverso contenuto di enzimi glicolitici e numero di mitocondri.
Le fibre ossidative sono più ricche di mitocondri e hanno una elevata capacità di produrre ATP con la fosforillazione ossidativa. Hanno però una bassa concentrazione di enzimi glicolitici.
Ogni muscolo del nostro corpo contiene entrambe le fibre, in proporzioni distinte.
Resistenza alla fatica e allenamento
La fatica è la condizione per cui a stimoli continui la forza espressa va via via calando.
Ci sono meccanismi differenti che la causano.
L'aumento di concentrazione di acido lattico intralcia il metabolismo del muscolo. Inoltre la contrazione dei vasi sanguigni diminuisce l'apporto di sangue che riesce ad arrivare ai muscoli. Inoltre può diminuire l'attività metabolica dei motoneuroni dopo frequenze molto alte di attività.
Nelle attività meno intense ma prolungate invece contribuisce molto la fine delle scorte di ATP.
Con l'allenamento si riesce a migliorare l'architettura cellulare del muscolo, ma anche la resistenza alla fatica del muscolo. L'allenamento può anche causare un rimodellamento del muscolo in base al tipo di attività per cui lo alleno (es° riconversione di fibre rapide glicolitiche in fibre rapide ossidative nell'esercizio aerobico).
Recettori muscolari per coordinare l'attività motoria
Ci son due tipi di muscoli che coordinano la attività muscolare
C'è il fuso neuromuscolare e l'organo tendineo del Golgi.
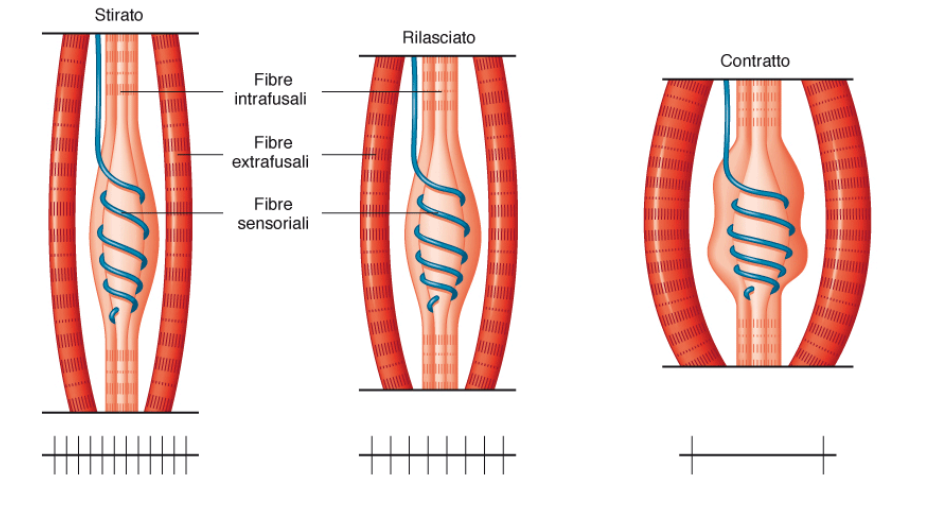
I fusi neuromuscolari sono fibre muscolari modificate, con una capsula di tessuto connettivo. Sono circondati da terminazioni sensoriali. Sono fibre muscolari di controllo che forniscono informazioni sullo stato di tensione di queste fibre. Lo stiramento dei muscoli attiva i recettori, in maniera proporzionale.
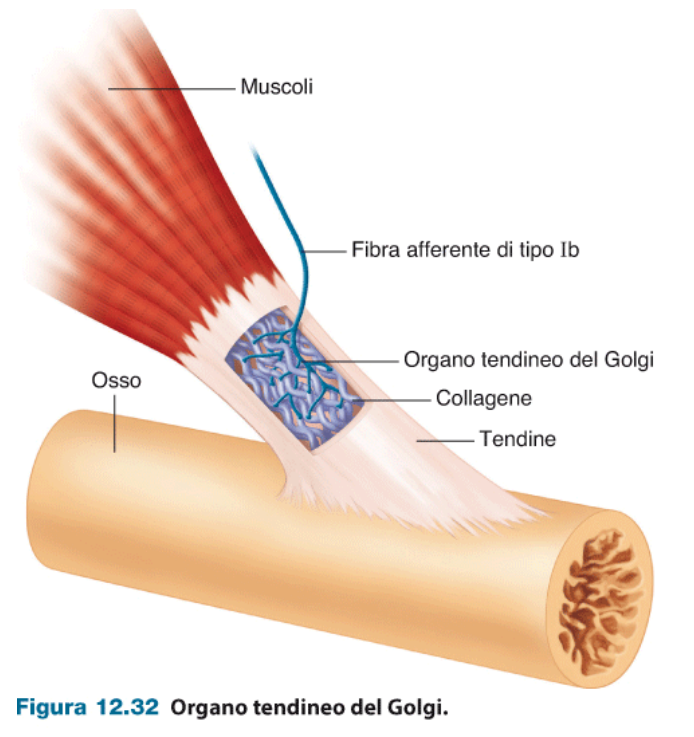
L'organo tendineo del Golgi fornisce più informazioni sullo stato di contrazione del muscolo. Misurano la tensione del muscolo, quindi misurano direttamente la forza che il muscolo sta esprimendo. Misurano quanto il tendine è stirato, la posizione del muscolo. Da questi segnali il cervello ottiene un feedback sullo stato di utilizzo del muscolo.
Questi organi hanno un effetto protettivo così da evitare un utilizzo eccessivo del muscolo tale che possa essere danneggiato.
Muscolo scheletrico, liscio e cardiaco
Nel nostro organismo esistono tre tipi diversi di muscoli.
I muscoli lisci hanno un'attività coordinata, perchè hanno delle giunzioni comunicanti che connettono gli stimoli elettrici. E' chiamato liscio perchè manca la striatura tipica dei muscoli scheletrici e cardiachi.
Tutte le strutture involontarie (organi interni, vasi sanguigni) hanno muscolatura liscia.
Nella muscolatura liscia non c'è un vero e proprio accorciamento, ma una contrazione della cellula, che diventa più piccola. La disposizione dei filamenti è obliqua.
Un'altra differenza è che la molecola che "blocca" il legame tra actina e miosina blocca la miosina invece che l'actina. Per il resto il ciclo dei ponti trasversali è invariato.
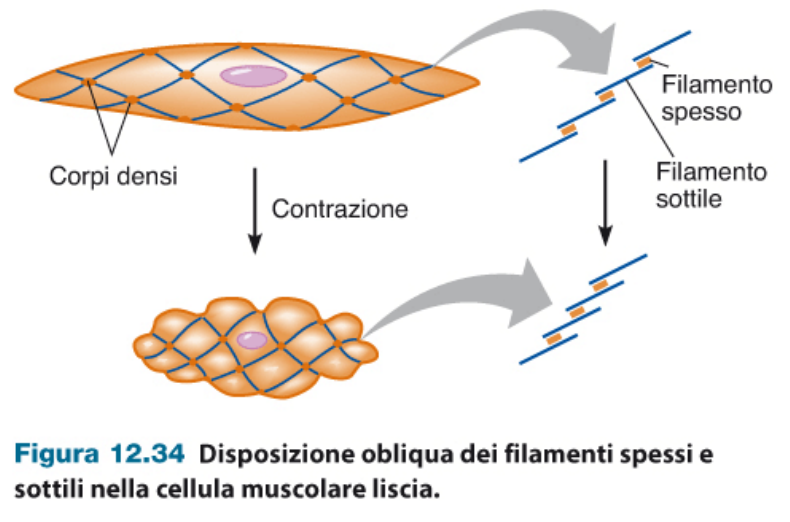
Mancano strutture come i tendini. Questa muscolatura è connessa direttamente a tessuto connettivo.